
Возможно ли повернуть вспять клеточное старение (сенесцент) и старение организма (сенилит) методами реювентализации и реювенилизации?
Статья - глава из книги-рукописи Гарбузова Г.А.: НЕОТЕНИЯ. ПРОДЛЕНИЕ МОЛОДОСТИ
Что такое онтофизис, его отличия от сенесцента и как он мешает реювенилизации у растений?
На сегодняшний день не известно ни одного случая полного омоложения из старика в юношу. В природе тоже такие факты не известны. Понятно специалисты на этот вопрос будут утверждать, что это пока недостижимо. Но это касается представителей животного мира. В растительном мире ситуация указывает, что такая реювенилизация* (на клеточном уровне) в принципе возможна.
Каковы механизмы реювенилизации у растений? Без реювенилизации на уровне меристем и побегов невозможна реювенилизация (на общеорганизменном уровне). У старых растений (деревьев) побеги существенно отличаются от таковых у молодых. Но эти качественные изменения связаны с онтофизисом*, а не с сенесцентом клеток, как это имеет место у животных. Это плацдарм, фундамент, с которого всё начинается. Под терминами реювенилизации (у растений) и реювентализации (у животных) автор понимает возврат к полной ювенильности клеток, полное восстановление тоти- и плюрипотентности. Тотипотентность позволяет восстановиться из одной клетки целиком всему организму, а плюрипотентность обеспечивает возможность к регенерации целых органов. Ни одной из этих способностей организм взрослого человека не обладает. Они попросту заблокированы полностью. Остаются открытыми только мультипотентные, унипотентные…, прогениторные уровни потенций клеток.
Особенность растений в том, что у них нет сенесцента клеток, но есть сенилит - старение, которое регулируется на общеорганизменном уровне. Это означает, что сенилит у растений регулируется только с одной стороны рычага маятниковой системы, то есть сверху (гормоны и др. профили), тогда как у животных маятник регулируется с двух сторон двумя рычагами: и сверху, и снизу (клеточные коммитации). У многолетних растений (деревьев) выражено проявление механизма возрастного онтофизиса*, то есть возрастного качественного изменения структуры тканей, в начале усиление, а на последних этапах жизни ослабление апикальных точек роста, что изменяет доминантные приоритеты, то есть соотношение роли камбиальных растущих тканей и апексов к стромальным (функциональным), смене баланса вегетативных побегов над генеративными. Это обуславливает особенности роста побега в дальнейшем, а также особой генеративной предрасположенности к гормонам. Это мешает тканям, побегам и клоновому потомству от взрослого и старого дерева вернуться в статус ювенильных.
- онтофизис базируется на структурно-гормональных перестройках на уровне тканей,
- сенесцент базируется на уровне внутриклеточных перестроек и образовании новых фенотипов.
Онтофизис и сенесцент – схожие процессы, ведущие к старению, но основаны на разных механизмах.
Онтофизис это закрепление неких возрастных (фенотипических на уровне эпигенома) и онтогенетических перестроек (морфотипических, на уровне генов), которые сохраняются и у линий клеток в культуре ткани, аналогично тому, как это сохранение происходит и у клоновых потомств в виде топофизиса (особенностей роста побегов: ортотропного /прямоствольность/ или плагиотропного /рост в бок/), или циклофизиса* (способность к закладке или не закладке тех или иных генеративных органов), или онтофизиса (с сохранением темпов клеточного роста или самообновления, либо их полной блокировки). Особенностью клеток животного типа является то, что до сих пор не найден универсальный на общеорганизменном уровне гормон реювентализации сенесцентных клеток. Ювентализация клеток здесь регулируется с нижнего клеточного уровня.
Особенно эти свойства возрастного онтофизиса сказываются на плохой способности к укоренению черенков (аналогия регенерации у животных) из кроны от старых деревьев. В тоже время черенки от молодых хвойных деревьев сохраняют некоторую способность к укоренению, но при этом часто проявляют топофизис, то есть кривизну и слабость роста у полученного клонового потомства из них. То есть, даже относительно молодые растения имеют ограниченный потенциал к регенерации. Тем не менее, в диссертационной работе по разработке ювенильных маточников для клонального размножения секвойи автор показал, что у неё возможно получить полностью тотипотентные клетки и побеги способные к 100% укореняемости и восстановлением к ювенильному морфотипу саженцев из них. См. рис. 1.
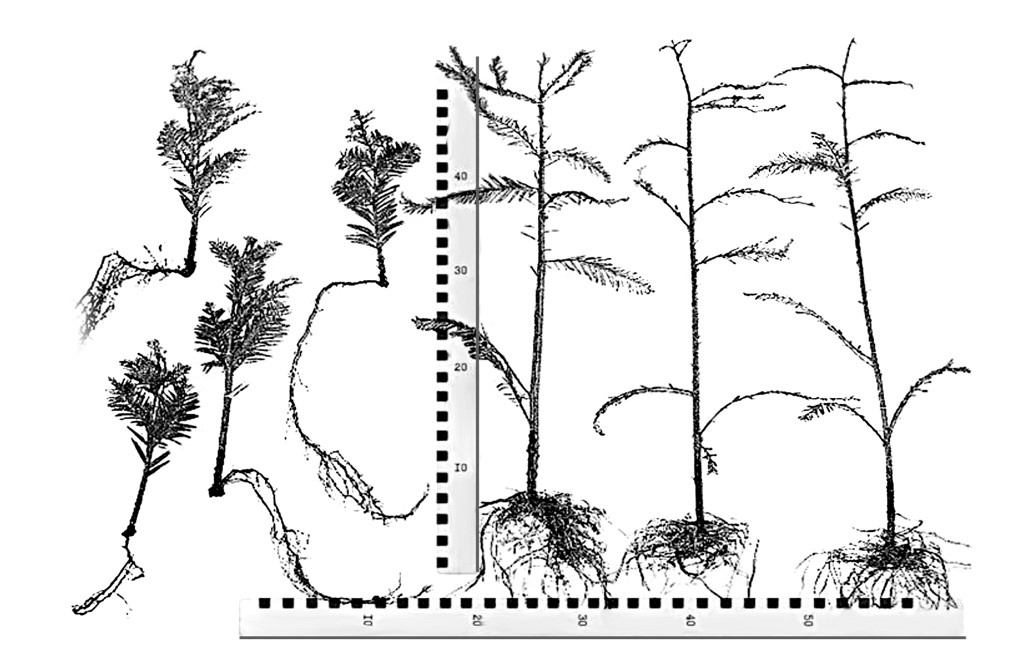
Рис. 1. Справа — внешний вид саженцев из укоренённых черенков от ювенильных ортотропных побегов, заготовленных в ювенильном маточнике, которые сохраняют все признаки ювенильного строения сеянцев и высокой степени к регенерации. Слева — укоренённые черенки от побегов, взятых в кроне.
Автор обнаружил у секвойи важную особенность – легко закладывать обилие поросли в основании даже старых деревьев, которые отрастали из спящих почек, обладающих высшей степенью ювенильности и тотипотентностью, см. рис. 2.

Рис. 2. На фото показано отрастание огромного количества ювенильных побегов порослевого типа, образующихся из спящих почек в основании ствола хвойного дерева секвойя вечнозелёная.
Отмечено, что такие спящие тотипотентные почки неограниченно могут закладываться-переоткладываться (мультиплицироваться) в зоне этиоляции побегов (см. рис. 3). Такое мультиплицирование может быть бесконечным, вплоть до многих тысяч лет. Это подтверждает, что клоновое воспроизводство имеет безлимитный потенциал, и что у растений нет внутренней клеточной программы на сенесцент, который, однако, имеется у животных.

Рис. 3. На фото представлены ювенильные побеги от порослевого ювенильного маточника секвойи.
Другой особенностью таких ювенильных этиолированных побегов является образование фасций – срастание побегов (см. на рис. 3 побеги под № 2, 4, 6, 9), что придаёт им необычный уродливый вид. Здесь чётко выраженное отсутствие апикального доминирования верхних почек, а доминирует нижняя корневая (ризомная) часть. Апикальное доминирование всегда ведёт к зрелости, а ризомное доминирование сохраняет ювенильность. Ризомная часть обеспечивает полную ювенильность (ювентальность), но только в сочетании с фактором отсутствия света (этиоляция). Именно в ризомной части вырабатывается основное количество гормона молодости - цитокинина. Это аналогично доминированию ювентальных клеток у животных над сенесцентными в детстве. Это говорит о том, что у животных тоже имеются ювентализирующие факторы, однако запускаются они опосредованно, через иммунную систему, которая и регулирует становление ювентальности-сенесцентности. Это особенность животного мира. Сенесцентность включается поэтапно через череду модификатов и фенотипов. И именно это этапное сенесцентирование аналогично действию апикального доминирования, которое у растений отключает закладку ювенильных почек, а у животных - сеть ювентальных стволовых клеток. В этих мезенхимальных точках роста и обновления происходит сенесцентное или ювентальное доминирование.
Это наглядный пример бесчисленной закладки спящих почек, из которых прорастают побеги с ювенильными свойствами. Причём нижняя часть этих вытянутых из-за темноты (находящихся под землёй) побегов этиолирована (не имеет окрашивания). Именно на этой этиолированной части особенно высока способность к бурной перезакладке на них новых спящих почек, и так до бесконечности. Это означает что бурно растущие побеги, но в темноте запускают обратный механизм – реювенилизации (снятие эффекта апикального доминирования)! Энергию роста спящим побегам даёт гормон цитокинин, который мощно вырабатывается корнями (ризома) и быстрее всего поступает к основанию дерева, где и находятся спящие почки.
Исходя из этого автор предложил технологию создания ювенильных черенковых маточников, см. рис. 4.

Рис. 4. Внешний вид порослевого маточника по технологии Г.А. Гарбузова с целью получения ортотропных ювенильных побегов для получения легко укореняемых черенков.
Важной особенностью этих маточников является получение неограниченного количества тотипотентных спящих почек и ювенильных побегов, причём с этиолированным основанием, см. рис. 5.

Рис. 5. Слева – черенки из кроны дерева. Справа - особенности черенков, заготавливаемых из ювенильных маточников с признаками этиоляции нижней части побега.
Отмечу что спящие тотипотентные почки и побеги образуются в основании дерева, возраст которого может исчисляться десятками-сотнями лет. Причём этиоляция (этиолирование) происходит в нижней части каждого ювенильного побега, которая находится ниже уровня земляного субстрата, то есть в темноте. Именно на этиолированной части побега закладываются настоящие спящие ювенильные почки, а из - них побеги ювенильного типа. Ювенильные побеги имеют отличающуюся внешние признаки и физиологические особенности, см. рис. 5 и рис. 6.
Итак, ювенильность и высокую потентность таким побегам обеспечивает:
- высокая энергия роста,
- гормон молодости – цитокинин,
- фактор темноты,
- …
Этот комплекс позволяет продлевать неопределённо долго статус ювенильности у растений.
Очевидно запуск подобного же комплекса факторов и мер должен способствовать продлению-восстановлению ювентальности и у клеток животных.
Идея создания ювенильных маточников у растений подтверждает возможность у них к возврату до полной изначальной степени потентности (через спящие этиолированные почки). Но возможны ли такие манипуляции у взрослого организма человека?
Особенности механизмов старения у животных. Сложностью является то, что механизмы старения в тканях животных имеют существенные отличия от растительного мира и регулируются одновременно на двух уровнях, где нижний этаж клеточный тоже имеет свои автономные механизмы сенесцента за счет программных коммитаций* и эпигенеза.
Причём, трофика и регулировка ювентальность/сенесцентность у клеток животных осуществляется не напрямую через гормоны, а опосредованно через иммунные клетки, действующих на клетки-бенефициары, а те на клеточном уровне выделяют свои регуляторы, а также свои SASP* или UASP*, которыми они и оперируют. SASP – это продукт стресса, который и определяет статус клетки. С повышением уровня клеточного стресса понижается степень-число ювентальных клеток.
В условиях культуры ткани, то есть изолирования клеток животного типа от условий организма, проявляется феномен лимита Хейфлика, что является одним из вариантов «вырывания» таких клеток из среды, определяющей статус ювентальности или сенесцента, то есть из общего контекста. Внешний фон созревания организма (его онтогенеза и прохождения по его этапам) имеет у клеток животного типа приоритетное значение для становления статуса сенесцентных или ювентальных клеток, которых нет у клеток растительного типа. Это является причиной неполноценности, то есть не полной автономности и ускоренной клеточной прогерии. Клетки попросту не могут переходить по ступеням фенотипии и ориентироваться по внешним факторам регулировок: вырваны из контекста. В условиях организмов животных изменены приоритеты важности сигналов регулировок идущих изнутри клеток, а затем после запуска интервенция сенесцента идёт и извне. От этих сигналов зависит быть клетке ювентальной или сенесцентной.
Особенности старения в растительном мире. Здесь нет клеточных барьеров сенесцента, что облегчает клоновое безлимитное существование даже в культуре клеток. У растений попросту не бывает фенотипа сенесцента, да ещё закрепляющегося на уровне эпигенома. Старение растений инициируется не с клеточного уровня, а общеорганизменного.

Рис. 6. На фото саженец Сосны Итальянской возрастом 2 года и имеющий выраженные особенности ювенильного фенотипа.
Показано, что свет, который попадает на верхушки побегов, является ингибитором ювенильности и фактором, который перезапускает в них механизмы выработки гормонов-ингибиторов (ауксина и др.) для спящих почек, но инициирующих механизмы дифференциации побега, выработки хлорофилла и закладки зрелых почек. По сути, эти почки на побегах являются аналогами сети очагов точек роста и самовозобновления из стволовых клеток, расположенных в каждой ткани у животных. И именно эта сеть стволовых клеток также теряет тотипотентность, затем плюрипотентность, а в конечном итоге даже и прогениторность, становясь сенесцентными.

Рис. 7. На фото побег из кроны взрослого дерева Сосны Итальянской, имеющий выраженные особые признаки зрелости, например, длинная жесткая хвоя собранная в пучки.
Удивительно то, что у растений часто можно встретить сохранение ювенильности у взрослых особей, см. рис. 8 и рис. 9.

Рис. 8. На фото изображен клоновый саженец от культивара «сорта» ели возрастом более 15 лет, побеги которого сохранили все признаки ювенильного типа охвоения, то есть проявили неотению. В норме такой тип ювенильного охвоения исчезает после 2 лет у сеянцев.

Рис. 9. Внешний вид ювенильной формы хвойного дерева ели (культивара с неотенией), на вершине которого выбились нормальные побеги типичные для взрослых растений. Возобладала нормальная меристема апикальных почек, в которой вернулась доминантность зрелости. Это указывает на лёгкую сохраняемость ювенильности у взрослых особей, то есть на продлённую неотению.
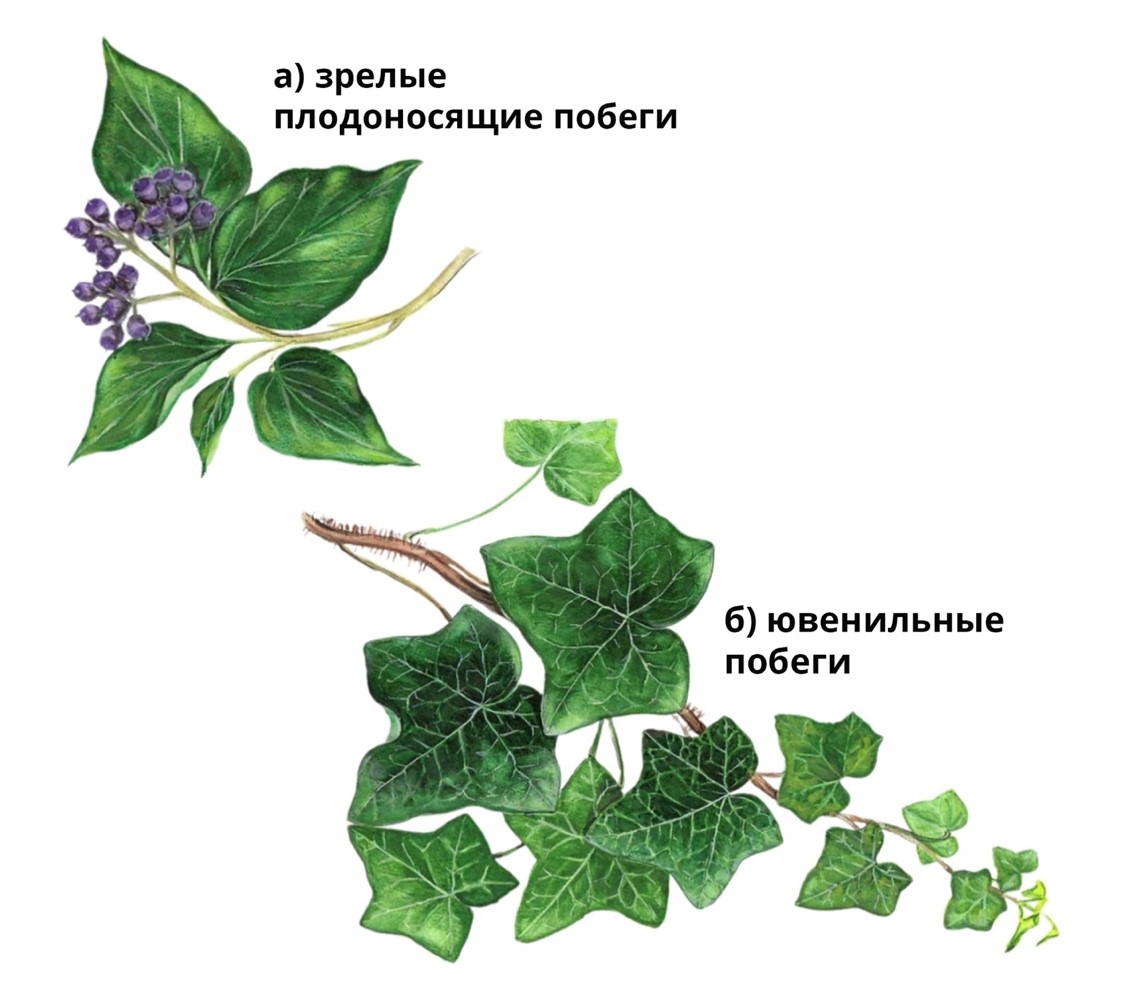
Рис. 10. а) взрослые плодоносящие побеги; б) ювенильные побеги плюща, взятые с одного растения. Фактором переключения с ювенильности на зрелость является степень освещенности данной части растения, а также степень концентрации ауксинов, которая зависит от мощи апикальной почек и общей биомассы растения. От света зависит доминирование в апикальных почках гормона зрелости - ауксина или цитокинина – гормона ювенильности. Растение легко переключается на неотению и может поддерживать её не ограниченно.
Следует отметить, что у растений существуют клоны-культивары, которые могут неограниченно поддерживать свою ювенильность и при этом это никак не отражается на их жизнеспособности. Так известен культивар криптомерии японской ювенильная форма, которая неограниченно сохраняет фенотип ювенильности, но при этом в шишках не образуются семена. Отсутствует мужская фертильность, но сохраняется женский потенциал. Очевидно повышен избыточно гормон ювенильности цитокинин, который и поддерживает фенотип ювенильности. См. рис. 11.
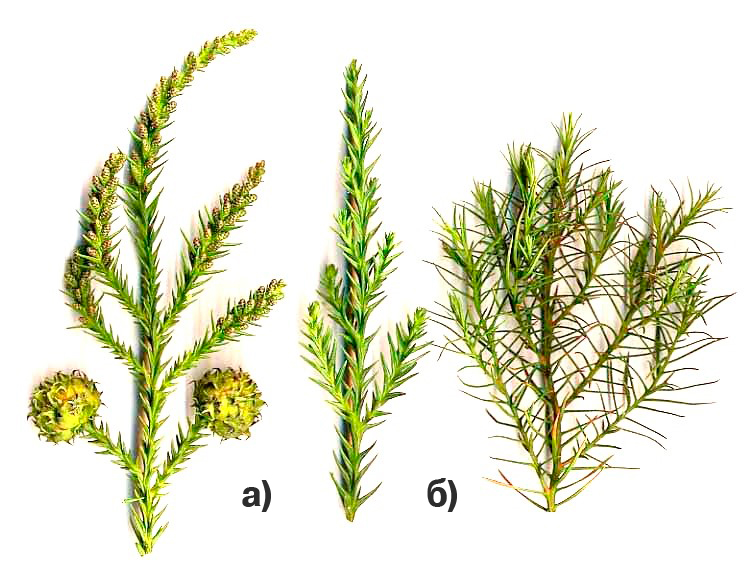
Рис. 11. а) обычные побеги криптомерии японской; б) побег от ювенильной формы, который способен закладывать репродуктивные женские стробилы, но не обладает мужской фертильностью, семена бесплодны. Черенки от ювенильных форм обладают высокой степенью регенерации (укореняемостью).
У растений, таким образом, зрелость или ювенильность зависят напрямую от уровня и соотношения гормонов (ауксины/цитокинины) и освещенности.
У животных нет гормона старения на общеорганизменном уровне, зато имеются автономные механизмы старения клеток, где и вырабатываются «гормоны» старения клеточного уровня. Таким образом нет такой прямолинейной зависимости, так как возрастная регулировка здесь дополнительно идёт и с другого этажа, то есть клеток, который обеспечивает механизм сенесцента – это специфика, которая появилась только у клеток животного типа и является дополнительным рычагом двухуровневой регулировки онтогенеза. У растений нет такого клеточного уровня регулировок, а онтогенез регулируется только с верхнего уровня общего гормонального фона. Другими словами, регулировка у растений происходит только с помощью одного рычага в маятнике. Это позволяет растениям легко переключать механизмы ювенильности и генеративной зрелости, то есть запускать реювенилизацию или сенилитированию своих побегов (статус за пределами зрелости). У растений также механизмами старения могут быть недостаток питания, света и гормонов или за счёт гормонов старения, например, абсцизовая кислота (АК). При этом АК может выступать в разных ролях: в одних случаях она:
- регулировщик апоптоза, например, для опадения листьев на зиму,
- регулировщик продолжительности жизни организма растения (1-, 2-х, 3-х-летки,
- регулировщик длительности покоя семян,
- регулировщик морфогенеза, в частности фасциации побегов (разъединения сросшихся побегов),
- …
Таким образом, видим, что один и тот же гормон может отвечать за разные предназначения. Очевидно, такую же роль играют и сенесцентные иммунные клетки:
- определяют созревание центральных «штабов» регулировок – гипоталамус, эпифиз, гипофиз, тимус,
- определяют модификаты и фенотипы иммунных клеток,
- ведут к запуску феноптоза* всего организма через механизмы анизомалий*,
- регулируют фасциации (например, перепонки на лапках у головастиков лягушек)
- регулировка морфогенеза у головастиков с рассасыванием хвоста,
- …
Примером регулировки зрелости за счёт только гормонов может быть растение плющ, где можно отметить, что побеги находящиеся в нижней части лианы (слабо освещенной), всегда проявляют ювенильность, а побеги в верхней части (находящиеся в условиях большей освещенности и имеют высокий уровень ауксинов) всегда проявляют генеративную зрелость, то есть обилие плодов, см. рис. 10. Это позволяет плющу безлимитный онтогенез! Есть растения, которые вегетативно в природе поддерживают своё существование десятки тысяч лет (известная самоклонирующаяся роща тополя осиноподобного [4]).
Влияния АК на продолжительность жизни и запуск старения на примере 2-х летнего растения сельдерей показан на рис. 12.

Рис. 12. На рис. видно: а) что переход растения сельдерей на второй год в стадию образования семян вызывает пожелтение и быстрое старение листьев, а затем отмирание; б) внешний вид растения на первый год жизни, когда не закладываются семена; имеет зелёный, молодой вид. Постоянное выщипывание семян (содержат гормон абсцизовую кислоту) переводит растение в 3-х, 4-х-летнее и т. д., сохраняет его неотеническую форму.
Однако, такого простого механизма в виде гормона старения у животных нет. Старение регулируется одновременно на разных этажах.
Неотения в мире животных. В свою очередь в животном мире тоже известны примеры пролонгированной или неограниченной ювенильности (неотения), но при этом животное может сохранять половую репродуктивную способность. Примером этого могут быть аксолотли, которые сохраняют фенотип ювенильности неограниченно, см. рис. 12 и рис. 13.
Неотения здесь связана с недоразвитием щитовидной железы. Она практически не синтезирует йод, который выполняет функцию активатора метаморфоза.
У аксолотля уровень тироксина недостаточен для трансформации (метаморфоза) в саламандру. Но щитовидка зависит от гипоталамуса, а его элевация связана триггерно с иммунной системой: пока предварительно в иммунной системе и эпифизе не произойдут необходимые перестройки гипоталамус тоже будет не зрелым. Аксолотли не проявляют признаков старения, которое реализуется через гиперзрелость (адальтус). Это особенность, связанная с неотенией — способностью достигать половой зрелости, не превратившись во взрослую форму (не претерпев метаморфоз). Функция генеративной зрелости здесь разъединена (не сопряжена) с функцией общей зрелости (метаморфоз). Аксолотли могут превращаться в амбистомы и во взрослую особь в условиях изменения среды в виде уменьшения влажности, пересыхания водоёма. По сути, это стресс, то есть здесь фактор стресса и запускает череду опосредующих нейро-иммунных механизмов, которые трансформируют гипоталамус. Это аналогично фактору света, который запускает генеративный морфогенез у растения плющ (рис. 10). В результате они утрачивают наружные жабры и переходят на обычное сухопутное дыхание. Также на это может повлиять простое добавление в пищу гормона щитовидной железы - тиреоидина. Так их можно "метаморфизировать" - превратить в амбистом. Пара месяцев замачивания мотыля перед кормлением в растворе тиреоидина позволят, чтобы аксолотли из водных существ приняли взрослую форму и далее жили на суше как саламандры.
Таким образом, устранив фактор стресса, у аксолотлей можно продлить неотению. Кстати, стресс как известно, значительно сокращает продолжительность жизни у человека. Но, тем не менее, стресс не является решающим фактором для продления жизни, так как работают еще и механизмы сенесцента, которые сопряжены с общим становлением, нормальным созреванием организма. То есть, нормальная реализация становления организма является тоже пусковым фактором для сенесцента.

Рис. 13. Аксолотль, ювенильная неотеническая форма.

Рис. 14. Взрослая форма саламандры.
У клеток животного типа тоже имеются ингибиторы и активаторы ювентальности, и активаторы и ингибиторы сенесцентности, а также другие специфичные факторы, действующие аналогично свету на растения и угнетающие ювентальность, но стимулируя степень зрелости и дифференциации. Аналогично регулировке зрелости через механизмы апикального или ризомного (корневого) доминирования у растений, у животных этот механизм регулируется через баланс доминантности ювентальных клеток или доминантности сенесцентных клеток. Причём первично эта регулировка зарождается и реализуется в иммунной системе, а затем передаётся на другие типы клеток и тканей [2]. Это означает что регулировки сенесцентности-ювентальности нужно осуществлять через иммунную систему!
Нейроны у землекопов (животное неотеник) сохраняют пластичность на протяжении всей жизни и ведут себя, как клетки новорождённых особей. Это указывает, что у нейронов здесь ограничен сенесцент.
О том, что сенесцент управляемый и наведённый процесс, а не процесс исходящий из клеточного лимита Хейфлика, говорят данные по особой, многократно раз большей продолжительности жизни у маток насекомых (пчёл, термитов). Матки живут в десятки раз дольше обычных термитов, даже при том что это особи одного вида. Значит онтодианная линия жизни* возможно у данного вида или у большинства видов может раздвигаться рядом факторов и не является строго предписанной. Так у маток термитов, несмотря на возраст, клетки сохраняют молодость (ювентальность). Но первична ли эта ювентальность или её спровоцировали верхние этажи? Очевидно, процесс в данном случае определяется нижним уровнем регулировок. Мощной поддержкой ювентальности клеток, а значит и ювентальности иммунитета, нервной и гормональной систем является то, что матки питаются маточным молочком, источника холина и фосфолипидов, а также содержит вителлогенин. Очевидно, это вещество экспрессирует специализированный для этого ген и повышает концентрацию ювентального (на уровне клеток) гормона в организме. У человека, очевидно, нет такого гена. Но ювентальность клеток есть и её определяют иммунные ювентальные клетки. Доминирование ювентальности сдвинуло сенилит в системе регулировок высших штабов. Сдерживание геронтоса* произошло не на уровне «штабов», а на уровне клеток (торможение сенесцента). Обеспечивается это работой двух особых генов, которые у маток-королев в 7 раз активнее, чем у рабочих особей. Один из них отвечает за борьбу с окислительным стрессом – именно он разрушает клетки и реализует старение. Второй регулирует клеточную коммуникацию. В результате клетки в возрасте 30 лет у королевы выглядят как у молодого организма. Окислительный стресс – это не просто стресс и сенесцент, а механизм регулировки созревания ткани или трансформации клеток.
Неотения у людей. Смотрите рис. 15.

Рис. 15. На фото 32-летний мужчина-неотеник, который перестал стареть 20 лет назад и выглядит как школьник.
Пример сохранения ювентальности и затем ювенильности среди людей?
Известен уникальный случай с мальчиком Денис Вашурин, когда из-за «генетических» нарушений и недостаточных возрастных перестроек в эпифизе (является драйвером гормонального созревания [4]) не смогли произойти возрастные перестройки в гипоталамусе (по причине отсутствия трансформации его иммунных клеток-резидентов из фенотипа ювентальности в фенотип сенесцента) и гипоталамус сохранил ювенильность своих функций. Врачи не смогли разобраться с истинной причиной данного случая и дать ему правильное объяснение. В диагнозе ему поставили гипофизарный нанизм – недоразвитие гипофиза.
Автор для объяснения предлагает здесь более конкретный механизм проблемы, так как, по сути, это следствие, а не причина. Гипофиз сам по себе не меняется, он зависим и созревает вместе с гипоталамусом, а гипоталамус зависит от динамики созревания и инволюции эпифиза. От мощи и доминирования эпифиза зависит продолжительность этапа юности, эта мощь у каждого вида своя и определяет продолжительность допубертатного этапа. Эту динамику в свою очередь определяет трансформация иммунных клеток эпифиза (их сенесцент подавляет активность функциональных клеток эпифиза). 40% клеток эпифиза это иммунные. Но запуск сенесцента иммунных клеток эпифиза зависит от динамики инволюции в тимусе. Через иммунитет происходит провоцирование сенесцента в эпифизе. Таким образом, нарушена целая цепочка последовательных изменений.
Автор видит, что начало этого процесса находится в отсутствии динамики появления новых возрастных иммунных фенотипов в макрофагах-резидентах (врождённый иммунитет). Не срабатывает какой-то сенсор-белок, рецептор, который является стартером и должен запустить цепочку (каскад) иммунных преобразований, ведущих к «созреванию» через механизмы таргетного регионального сенесцента или апоптоза именно в эпифизе. В частности, угасание эпифиза оказывает противофазное активирование со стороны гипоталамуса, то есть ведёт к созреванию = растормаживанию гипоталамуса. У детей эпифиз имеет большие размеры и высокую функциональную активность. Несбалансированный (митозами) апоптоз и сенесцент имеют много общего, и возрастная трансформация эпифиза происходит за счёт этих механизмов. В эпифизе нарастают дистрофические процессы, увеличивается метавоспаление, которое вызывает кальциноз железы… Железу грубо отключают из общей цепи регулировок вегетативных и трофических функций, сохраняя только минимальный контроль за циркадными процессами.
Таким образом через таргетные механизмы сенесцента регулируются:
- на этапе юности механизмы сенесцента и апоптоза реализуются (открываются) не на уровне всего организма (глобально), а сугубо локально, направленно, таргетно, благодаря аффинности иммунных клеток эпифиза к сигналингу из тимуса,
- первично программа сенесцента зарождается в тимусе благодаря кейлонным* механизмам самосозревания; это является началом для запуска механизмов трансформинга (разворачивания) в других «штабах» регулировок, включая эпифиз, гипоталамус…
- эпифиз является перекрёстком взаимодействия иммунонейрогуморальных регулировок, включая становление этапов онтогенеза,
- гипоталамус является перекрёстком нейрогормональных регулировок, включая становление этапов онтогенеза.
- через взаимодействие этих штабов (эпифиз – гипоталамус) происходит состыковка, синхронизация работы в различных «штабах» систем организма: гормональной, иммунной, нейрогенной…,
- доминирование одного из штабов определяет возрастную динамику, зрелость, переходы по этапам онтогенеза,
- до 12 лет проявляется эпифизарное доминирование и эпифизарная молодость, а после 12 лет проявляется гипоталамическое доминирование и гипоталамическая зрелость с коррекцией в репродуктивную зрелость и торможения вегетативных функций (рост),
- динамика трансформаций в гипоталамусе после 12 лет не останавливается, а нарастает до 40 лет, что включает механизмы гиперзрелости = климакса и переход на новый этап онтогенеза – матура, а затем – сенилит, то есть инволюций в штабах и дистрофий на местах.
Как вывод:
- всеобщий сенесцент реализуется первично через сеть сенесцента в регуляторах развития = «штабах», что проявляется в их разной трансформации в разных отделах: в виде элевации или инволюции;
- действие медиаторов-гормонов эпифиза имеют сродство (аффинность к рецепторам) преимущественно на гипоталамус, а не на весь организм;
- снижение активности эпифиза растормаживает гипоталамус, что ведёт к повышению уровня гормонального фона, зрелости, перезрелости, а затем и сенилита (адальтус, матура, сенилит);
- повышение активности (функциональности) эпифиза наоборот подавляет (инактивирует) гипоталамус, что ведёт к снижению чрезмерного гормонального фона, перебалансировке его состава и возврата в исходное юное состояние;
- эпифиз обеспечивает статус «эпифизарной молодости» до 12 лет, сдерживая созревание гипоталамуса;
- эти штабы-центры (эпифиз-гипоталамус) являются «воронкой» (пропускным каналом) для управляемого, а затем интервенционного растекания сенесцента из точек его зарождения в иммунной системе;
- эпифиз одновременно является драйвером онтогенеза: на определённой высоте его активности (доминантности) он поддерживает статус ювенильности, а при отсутствии доминантности – открывает путь к зрелости, гиперзрелости и сенилиту;
- через реювенилизацию эпифиза (пропускная воронка) можно поддерживать неотению и остановить цепную тотальную реакцию сенесцента (интервенцию) на весь организм;
- если удастся сохранить или реювенилизировать эпифиз, то это позволит достигнуть неотении;
- воздействуя на иммунную систему «штабов» можно корректировать статус их зрелости.
Важно получить ответ, что будет если реювенилизировать эпифиз в зрелом возрасте (матура) или сенилите? Поддержка ювенильности штабов на этапе юности поможет пролонгировать молодость и проявить неотению. На других этапах онтогенеза наблюдается тотальный процесс инволюций в штабах и дистрофий на местах. Допустим мы усилим поставку мелатонина в организм в 10 раз (как в юности), то это не означает, что нам удастся разбудить инволютированный гипоталамус, так как в нём произошли дистрофические процессы и высокая степень резистентности. Сигналинг из эпифиза пойдёт в никуда, так как мишени резистентны. Вслед за гипоталамусом должны ювенилизироваться ткани-мишени на местах, но там тоже высочайшая степень резистентности и дистрофий. Это означает, что реювенилизацию одновременно надо проводить не только на уровне штабов, но и на нижних уровнях клеток и тканей. Очевидно, для этого надо будет подключать механизмы парабиогенезной реювентализации и CAR-T технологий по переобучению иммунной системы, а также аутофагию, полуавтономии и факторы нейрогенной коррекции, включая световую депривацию на длительный период (циклами на 7 дней) и...
Предназначение адаптивного (приобретённого) иммунитета в тотальном распостранении сенесцента. Тем не менее, есть специалисты, которые связывают сенесцент с адаптивным иммунитетом (приобретённый в процессе жизни), который основывается на работе Т- и В-клеток, — единственном типе клеток, внутри которых происходят перестройки и изменения ДНК в некоторых генах. В тимусе происходит обучение Т- и В-клеток на трансформацию резидентов, утрате ими ювентальных свойств. В других типах клеток в тканях и органах этого нет! Скорее всего, врождённые макрофаги-резиденты тимуса кейлонным* путём первично запускают свою активность в тимусе, и побуждает к срабатыванию, реагированию, обучает на это приобретённый иммунитет, который по сути является фактором сигналинга, что и распространяет наученный сигнал дальше. Дальше обученные мобильные Т- и В-клетки срабатывают в иммунных клетках-резидентах эпифиза и трансформируют его. В итоге повышается его ризистентность и понижается функциональность. Угасание эпифиза растормаживает гипоталамус, который активирует гипофиз. Слабым местом этой концепции является отсутствие объяснения, как обучаются первично клетки-учители, откуда они получают «инструктаж», сигналинг для распространения. Автор сторонник кейлонного зарождения сенесцента в тимусе.
Возрастная фенотипия и является тем драйвером, который ведёт к созреванию в «штабах» регулировок, а затем распространяется в виде «созревания» через модификаты, но, по сути, ведущие к их сенесценту и затем тканей всего организма. В обсуждаемом случае неотении у мальчика-мужчины, очевидно, первично и произошло такое нарушение с сохранением ювентальности у иммунных клеток-резидентов, что не позволило сработать иммунным драйверам зрелости и угнетения ювентального фенотипа иммунных клеток.
Похоже такое сохранение ювентальности клеток у людей является аналогией, продлённой ювенильности (неотении) и у взрослых деревьев, то есть сбоем регулировок, которое обсуждалось выше. Причём у растений эта пролонгированная ювенильность может быть не обязательно результатом сбоя неких генов, контролирующих это, а управляемым процессом, и, регулируемым рядом внешних факторов и гормонами, см. рис. 10. Как видим, ювенильность у растений легко регулируется гормонами (соотношением баланса цитокининов и ауксинов).
При возрастных перестройках эпифиза затрагиваются не только иммунные клетки, но и стем-клетки. Типичное старение-сенесцент в данной ткани, как это происходит и в любой ткани. При этом в клетках происходит перераспределение в соотношении различных путей энергетического обеспечения: начинает преобладать анаэробный гликолиз, провоспаление, оксидативный стресс, дистрофия...
Без освобождения от тормозящих сигналов из эпифиза (из-за его инволюции) гипоталамус не смог поднять свой уровень гормональных регулировок до возрастной нормы зрелости и преодолеть этап пубертата. Зрелость гипоталамуса в свою очередь усиливает инволюцию эпифиза. При этом мужчина в возрасте 32 года выглядел внешне как 14-летний школьник.
Причина проявления неотении в данном случае явно находится в проблеме иммунных клеток-резидентов к способности перестраиваться в новые возрастные фенотипы. Речь идёт о трансформации в новые возрастные фенотипы типа модификатов М1 и М2. Но эти модификаты являются результатом появления митохондриальных модификатов ММ1, ММ2.
Фенотипы, модификаты, сенесцент тоже реализуются через эпигенез и тоже сцеплены с тем же блоком генов. В отличие от обычных иммунных фенотипов (М1, М2…), при сенесценте эта фенотипия переходит в устойчивую модификацию (невозвратную, то есть изменения происходят в большей степени не со стороны функциональных возможностей, а структурно на уровне энергетики и митохондрий).
Отключен механизм (ген) коммитации? В результате в иммунной системе тимуса (или гипоталамусе) утрачена способность модифицироваться из ювентальных в сенесцентные клетки. Без этого нейроиммунные клетки гипоталамуса не могут в достаточной степени получить сигнал на дальнейшее созревание и продвигаться по следующим ступеням дифференциации, и остаются в недоразвитом состоянии. Как итог, у этого человека сдержан метаморфоз, точнее осуществлён неполный метаморфоз, когда фенотип юности переходит в фенотип адальтуса. Некоторые элементы метаморфоза осуществились, так как ограничен рост – гомойотопия*. При этом разорвана цепочка событий на клеточном уровне для последовательной реализации сенесцента в новых клеточных локализациях. Разрыв этой цепочки может отобразиться на последовательности событий на общеорганизменном уровне и гормонов, а значит смены (отмены) механизмов сенилита. Сенесцент определяет уровень и темпы сенилита, а не наоборот. Только на конечных этапах онтогенеза, когда происходят дегенеративные процессы, сенилит способствует сенесценту. Сенилит базируется и определяется сенесцентом. А точнее от степени сенесцентирования (сенесцентной доминантности) зависит и продвижение по этапам онтогенеза, включая геронтос. Геронтос = сенесцент + сенилит. Это означает, что старение у человека вписано в онтогенез на клеточном уровне и задаётся (исходит) снизу, а не формируется на верхних этажах регулировок. Очевидно на клеточном уровне цепочка последовательных переходов клеточной ювентальной доминанты в сенесцентную доминанту у этого человека тоже будет ограничена. Если это так, то данная неотения обеспечит меньшее проявление у него проблем старости в виде анизомалий* и появления старческих фенотипов (облика).
Очевидно в случае с этим мальчиком произошел эффект резистентности (утрата чувствительности) иммунных клеток к образованию новых модификатов (в т.ч. и сенесцентных), а те в свою очередь потеряли способность вести к перестроечным процессам (через инволютивные механизмы) в штабах регулировок и тем самым переключаться на новые этапы онтогенеза.
Сдерживание роста связано с гормоном роста, выработку которого инициирует гипоталамус. Но такой линейный рост возможен в норме только до 12 лет, затем, когда созревает половая система, происходит окостенение костных эпифизов, закрывающих дальнейший рост, но уровень гормона роста мало снижается.
Очевидно следует отличать неполный метаморфоз от полного отсутствия метаморфоза, который будет также сопровождаться не только отсутствием генеративной зрелости, но и ограничением роста (гомойотопия*), см. рис. 16.

Рис. 16. Пример неотении у взрослого человека с остановкой роста (гомойотопия).
Таким образом, очевидно, произошло ровным счётом всё наоборот тому, что происходит при прогерии. Если при прогерии происходит ускоренный клеточный сенесцент, то при неотении – остановка клеточного сенесцента или пролонгирование клеточной ювентальности.
Что общего и какие различия у сенесцентных и онкологических клеток?
По своей сути это аберрантные клетки, которые вышли за пределы гомеостаза. Это было показано автором [2, 3]. Но механизмы их проявления и генная база реализации общая. Это важно понимать, так, как это указывает на схожесть методик их преодоления, а также поиск этих методик. В обоих случаях у них происходит:
- вход в гликолиз,
- отключение защитного апоптоза,
- аберрантная аутофагия (парадоксальная),
- запуск механизмов провоспаления,
- слом механизма репликаций,
- нарушение резистентности, чувствительности к сигналам,
- ...
Обращает внимание факт, что при сенесценте и онкологии запускается единая система реагирования путем выработки провоспалительных цитокинов. Но оказывается и в этой единой маятниковой системе имеются разные крылья регулирования. Это даёт различия в эффекте провоспалительных цитокинов при онкологии и сенесцентном старении клеток. При онкологии цитокины, свойственные «гуморальному» иммунному ответу (Th2-тип), стимулируют развитие опухоли. Th2-тип иммунного ответа (иммунный ответ 2-го типа, Т2-воспаление) — это тип иммунного ответа, при котором активация Т-хелперов 2-го типа (Th2-клеток) приводит к развитию гуморального иммунного ответа. Уточняю что Th2-тип это приобретённый специфический иммунитет, так как он обеспечивается работой Т-хелперов 2-го типа (Th2) — субпопуляции Т-лимфоцитов. Это позволяет уточнить, увидеть в чём разница, что онкологическое провоспаление активируется специфическим иммунитетом, тогда как сенесцентное провоспаление регулируется Th1-тип иммунного ответа — это неспецифический врождённый иммунитет, который обеспечивает клеточный иммунный ответ. Врожденный – приобретённый; специфический – неспецифический иммунитет – это два крыла единой маятниковой системы. Сенесцент запускается через врожденный иммунитет. Именно этот иммунитет в первую очередь реагирует на возрастную динамику гормонального фона, воспринимая его на определённом этапе как стресс (выход за пределы клиренса гомеостазов) и реагируя в виде сенесцента.
Ключевое слово здесь: стресс на возрастную динамику. Это драйвер сенесцента. Устранить возрастную динамику, зрелость нам не под силу, то есть сохранять неотению*. Но ослабить эффект этой возрастной динамики на сенесцент можно путём интервального голодания, которое ограничивает провоспалительное проявление у сенесцентных клеток, устраняет возрастную резистентность клеток, например, инсулинорезистентность. Некоторые исследования показывают, что у людей, придерживающихся различных режимов интервального голодания, отмечается снижение уровня триглицеридов и «плохого» холестерина, а также повышение уровня «хорошего» холестерина. Но ограничение провоспаление можно достигнуть только в пределах узких рамок клиренса, выход за его пределы поведет к усилению провоспаления и сенесцента, а шлейфом пойдут инсулинорезистенность, холестериновые проблемы и огромный спектр других показателей.
Стрессом для клеток также может быть фактор отсутствия взаимодействия, единства с организмом (эффект лимита Хейфлика). Это связано с механизмами двух-ярусных регулировок (зависимость клеток и организма), тогда как у растений это одноярусный тип регулировок. Стресс на возрастную динамику, а также на отсутствие единства с организмом реализуется через врождённую иммунную систему. Очевидно интервальное голодание в культуре ткани тоже ослабит сенесцент, т.е. лимит Хейфлика.
Не удивительно, что цитокины «клеточного иммунного ответа» (Th1-тип), наоборот, способны подавлять опухоль. Цитокиновый состав сенесцентных и опухолевых провоспалительных молекул несколько разный.
В опухолевых клетках происходит продукция провоспалительных цитокинов, среди которых IL-1, TNF-α, IL-6.
При сенесцентном старении провоспалительные цитокины выбрасываются локально в начале самими иммунными сенесцентными клетками (макрофагами-резидентами), которые, несмотря на отсутствие репликативной активности, влияют на клетки-соседей, в том числе и на клетки-бенефициары, кроме того, они активируют дрейфующие, подвижные клетки приобретённой иммунной системы и вызывают более широкий фронт хронического генерализованного воспаления. В качестве маркеров сенесценции используются, например, IL-6 и IL-8.
Таким образом, эффекты цитокинов при онкологии и сенесцентном старении отличаются: при онкологии одни цитокины стимулируют развитие опухоли, другие, наоборот, её подавляют, а при сенесцентном старении провоспалительные цитокины сами вызывают воспаление.
Особенность онкологических клеток в том, что ряд триггерных факторов (специфического характера) уводит их в стресс (через иммунитет Th2), но в более глубокую одностороннюю крайность (за рамки гомеостаза) - в сторону полной разблокировки репликаций, то есть резко сдвинут баланс генного механизма регулировок репликаций между транспозитными и контрпозитными рычагами авторегулировок. Диапазон возможностей адекватного реагирования полностью сужен и смещен в одну сторону. Это происходит за счет образования эпигенетических модификаций из-за анизопаттернов* и энтропостаза*, что ведёт к блокировке-отключению (репрессии) тормозных генов, отвечающих за своевременное торможение репликаций, представляющих собой механизм обратного выхода из репликаций и играющего роль контрпозитного рычага. Репликации должны своевременно включаться (транспозит) и своевременно отключаться (контрпозит).
В свою очередь у сенесцентных клеток этот же механизм регулировок репликаций в процессе эпистресса* с последующим эпигенезом (смещением) уходит наоборот в противоположную сторону в крайнюю её часть – полной остановки репликаций. Сенесцентный эпистресс в результате смещения возрастного фона действует мягко и на него настроены только макрофаги-резиденты. Поэтому сенесцент это стресс среди резидентных иммунных клеток.
Но в обоих случаях их механизмы связаны с сочетанием своих факторов неспецифического и специфического стресса, причём при онкологии это свои специфические факторы, которые отличаются от специфических факторов при сенесценте. При этом неспецифические факторы (механизмы) стресса у них едины и ведут к образованию УКАМ* (основа для сенесцента), отличаются только специфические, что ведёт к образованию УКОМ* – универсального клеточного онкологического модуса = онкотрансформата. Итак, особое сочетание специфических и неспецифических факторов определяет направленность трансформации в две крайности: сенесцентность или онкологичность. Эти две крайности всегда работают за пределами той или иной стороны (транс- и контрпозиты) единого гомеостаза репликаций (анизостаз*, а точнее трансанизостаз* и контранизостаз*). Онкология – это контранизостаз, сенесцент – это трансанизостаз. Сенесцентные и онкологические модусы у каждого морфотипа клеток проявляется по-своему, что образует широкий спектр их разнообразия. Таким образом при онкотрансформации нарушаются, добавляются (включаются, усиливаются «взламываются» генные программы), которые являются специфичными триггерами и способны перетягивать канат клеточного стресса в другую сторону. На эпигеноме это фиксируется, закрепляется и в итоге образуется новый устойчивый модификат (фенотип).
Наука вышла на передовые рубежи по разработке методов уничтожения раковых клеток с помощью иммунитета. Достигнуты положительные результаты при помощи CAR-T технологии (переобучение иммунитета). Но это же является подсказкой, что также можно бороться и с сенесцентными клетками!
У каждого типа сенесцентных или онкологических клеток имеется свой комплекс сочетания специфичных и неспецифичных факторов стресса нужных для их проявления. Это проявление всегда осуществляется через специфические белки их реализации. Каждый белок имеет свои гены для его считывания. CAR-T терапия направлена на коррекцию именно этих белков и их генов.
Особенностью сенесцентных клеток является то, что специфическим и неспецифическим стрессом для них являются три фактора:
- Динамика возрастных неизбежных сдвигов на уровне всего организма [1] (специфический фактор).
- Динамика возрастных неизбежных сдвигов на клеточном уровне (модификаты, фенотипия за счёт эпигенеза), в частности нарастание доминирования сенесцентных клеток, появления среди клеток новых модификатов, как промежуточных форм для появления устойчивых фенотипов.
- Динамика взаимодействия факторов №1 и №2 (смена значимости), то есть как с факторами на уровне всего организма, так и интервенционным влиянием со стороны других стареющих тканей через SASP* (неспецифический фактор) [4].
На фактор №2 можно воздействовать только через метод полуавтономного циклического питания-голодания и усиления аутофагии. Напомню, что у растений найден один из факторов (этиоляция пробуждает только спящие-ювенильные почки), который запускает, активирует гены ювенильности и плюрипотентности, а значит отвечающие за полную ювенилизацию. У клонового потомства будет полное обновление онтогенетического цикла! Естественно, такие гены ювенильности есть и в животном мире, а значит следует ожидать, что можно найти факторы, которые разблокируют их.
Похоже, что полуавтономия будет воздействовать на организм, стимулируя в нём запуск ювентальных клеток (аналогично этиоляции у растений), а также перестраивая верхние этажи регулировок на уровень юности.
Подробно о методе полуавтономии в книге [2] и в моей книге-рукописи: «Неотения. Продление молодости».
Что такое CAR-T технологии и их возможности воздействовать на нижний клеточный уровень №1?
Они действуют через иммунитет, его переобучение и могут повлиять только на сенесцентные и иммунные клетки, то есть на нижний клеточный уровень №1 (неспецифический), но не влияют на верхний уровень (эпифиз…, т.е. специфический). Таким образом, это односторонний подход, который может решить задачу на одном уровне, но который не убирает главную причину: возрастную динамику перестроек. Да возрастные взаимные перестройки зависят от каждого уровня №1 и №2, но у каждого уровня есть своя автономия, клиренс (зазор) свободы действия, независящий от противоположного уровня. Этот клиренс позволяет каждому уровню самостоятельно «наполниться», созреть и только при достижении порога повлиять на противоположный. Благодаря этому клиренсу происходит ступенчатое движение-проявление онтогенеза.
Отмечу, что CAR-T технологии могут по сути заменить или усилить метод сенолитиков*, которые усиливают выбраковку сенесцентных клеток и открывают дорогу ювентальным. Потенциал метода сенолитиков крайне ограничен и вряд ли сможет решить эту проблему на широком уровне, то есть в полном объёме.
Тем не менее, CAR-T технологии позволили получить успех у 60% онкопациентов. Феноменальный результат при миеломной болезни: в этом конкретном случае констатируют 100% эффективность. Метод CAR-T (Chimeric Antigen Receptor T-cells) – это сложнейшая биотехнологическая процедура, которую можно назвать «персонализированным лекарством».
Забор клеток. У пациента забирается кровь, из которой методом афереза выделяются собственные иммунные клетки – T-лимфоциты.
Генетическая модификация. Это ключевой этап. В лабораторных условиях с помощью обезвреженного вируса-переносчика гены T-лимфоцитов изменяют. Им «встраивают» специальный химерный антигенный рецептор, который запрограммирован на точное распознавание белка на поверхности опухолевых клеток.
Культивация. Генетически «перепрограммированные» клетки размножаются в искусственных условиях до количества в десятки и даже сотни миллионов экземпляров.
Возвращение в организм. Полученную армию «киллеров» вводят обратно пациенту. Эти клетки теперь умеют находить, атаковать и уничтожать рак, запуская мощнейшую и прицельную иммунную реакцию.
Оказывается, эту же технологию разрабатывают и для лечения клеточного сенесцента! Здесь тоже иммунные клетки модифицируют так, чтобы они распознавали распространённый на поверхности сенесцентных клеток белок. В результате метод оказался эффективен и улучшил состояние мышей при фиброзе лёгких и печени, а также при стеатогепатите.
Однако стратегии борьбы со старением с использованием CAR-T-клеточных препаратов требуют дальнейшего изучения. Возможны иммунные осложнения, кроме того, уничтожение сенесцентных клеток с использованием данного класса препаратов сложно остановить при необходимости.
Интервальное голодание с ограничением калорийности на фоне парабиогенных приёмов с целью реювентализации на клеточном уровне
После 16 часов голода у клеток начинает ослабляться сенесцентная резистентность, например, в мышцах и печени вдвое увеличивается количество рецепторов к инсулину, уходит инсулинорезистентность. Организм не просто меньше производит инсулина — он учится слышать его снова, что критически важно при инсулинорезистентности.
Основная роль гормона роста в период голода — активация аутофагии («самопоедания» повреждённых клеток) и липолиза (расщепления жира). Он работает как метаболический мусорщик, а не культурист.
Кратковременное повышение кортизола утром (на 12-16 часу голода) имеет адаптивный характер. Он мобилизует жиры, не вызывая катаболизма мышц, если вы не голодаете сутками. Это гигиенический стресс, как контрастный душ для метаболизма.
При голодании 16/8 уровень гормона адипонектин, вырабатываемого жировой тканью, увеличивается на 20-30%. Адипонектин повышает чувствительность к инсулину и напрямую заставляет митохондрии сжигать жир для получения энергии. Тело переключается в режим эффективного жиросжигания даже в состоянии покоя.
При схеме 16/8 скорость основного обмена не снижается, а в некоторых случаях увеличивается на 3-5% за счёт:
Повышения термогенеза (выработки тепла).
Увеличения активности бурого жира.
Кратковременное голодание может улучшать когнитивные функции, активировать мозг и даже защищать его от старения. Голод стимулирует выработку нейротрофического фактора мозга (BDNF) – вещества, которое помогает создавать новые нейронные связи, улучшает память и защищает от возрастных когнитивных нарушений. Активируются кетоны – альтернативный источник энергии для мозга, который работает эффективнее глюкозы.
Снижается уровень воспалений, связанных с болезнью Альцгеймера и деменцией. Доказано, что интервальное голодание увеличивает количество митохондрий в клетках мозга, что делает его более устойчивым к стрессу и улучшает когнитивные функции.
Термины
- Анизопаттерны — патологически изменённые — паттерны на эпигеноме, то есть противоположноть норма-паттернам у здоровых ювентальных клеток. — Термин предложил Г. А. Гарбузов.
- Геронтос — общее старение регулируется одновременно двумя рычагами на двух этажах: на нижнем уровне фенотипа клеток (сенесцент) и на верхнем уровне всего организма в виде онтофизисных фенотипов (сенилит). — Термин предложил Г. А. Гарбузов.
- Гомойотопия — ограниченность роста, от слова «гомойо» — способность всех особей вида или органов в них достигать жёстко заданных величин, например, размеров, а «топо» — от слова «топология» — размеры, пространство. — Термин предложил Г. А. Гарбузов.
- Кейлонный механизм регулировки — первично инициирующее программируемое изнутри клетки звено регуляции клеточного гомеостаза тканей, которое происходит за счёт перестройки и изменения ДНК в некоторых генах и обеспечивающее контроль над процессами модифкаций (или фенотипов, или субпопуляций) и размножения клеток.
- Контранизостаз — это выход функции репликаций клетки за пределы репликативного гомеостаза и вход в анизостаз онкологической направленности. — Термин предложил Г.А. Гарбузов.
- Онтофизис — у растений подразумевает не простое закрепление у клонов неких особенностей роста и плодоношения, а проявление более общего процесса, то есть сохранения степени онтостадийной зрелости, в том числе и высшей её степени, когда полностью заблокированы рост или плодоношение на нижних побегах. Но блокировка происходит не на уровне генов, а на уровне сложившегося комплекса активности камбиальной ткани и соответствующей ей стромы. В отличие от высшей степени проявления онтофизиса у побегов растений, в животном мире аналогом этому является сенесцент у клеток. Онтофизис = сенесценту. Разблокировать такое состояние у растений возможно только при особых условиях реювенилизации, то есть снятии апикального доминирования, и обязательном превалировании цитокининового фона. При этом расширено понимание данного явления не только для клонов, но и в пределах данной особи растения, когда новое поколение побегов в пределах одного и того же дерева проявляет продвинутую физиологическую или морфологическую стадийную зрелость. Программы на ювенильность у них заблокированы.
- Онтофизис в животном мире подразумевает образование множественного разнообразия возрастных фенотипов клеток. Это в свою очередь откладывает свой отпечаток, отражение на возрастных этапах в виде качественных изменений во всех структурах организма. Фенотипия (не путать с морфотипами) клеток определяет онтофизис (сенесцент) на уровне тканей. Набор определённой критической массы переходов по фенотипическим ступеням определяет становление сенесцента. Причём такая возможность сенесцента заранее предусмотрена эволюционно для клеток животных. Это и есть механизм коммитации, то есть перехода на сенесцент. Кроме морфотипической динамики в тканях и органах с возрастом происходят (накладываются) онтофизисные процессы. Они основаны на динамике соотношения ювентальных/сенесцентных клеток, доминирования того или иного типа их фенотипов. Морфо- и фенотипы в совокупности в масштабах всего организма отображают этапы онтогенеза (детство, юность, адальтус, матура, сенилит). При этом фенотипы клеток отображают изменчивые модификаты и трансформаты для некоторых морфотипов. Фенотипы накладываются на морфотипы, которые могут быть представлены рядом фенотипов. Точнее, когда фенотипы сенесцента становятся доминирующими над ювентальными, они запускают усиление проявления фенотипов онтофизиса в тканях и органах; в том числе это является причиной возрастных перестроек в интегрирующих всё «штабах» регулировок, включая элевацию в одних и инволюцию в других. Это и является механизмом реализации становления (через ступени на онтодианной линии). При пересадке старых органов молодым особям это их состояние старости отражается на организме-акцепторе. У животных регулировка запускается через статус клеток иммунной системы, уровень их трансформации в возрастные фенотипы. Это обеспечивает механизм доминантности ювентальности/сенесцентности. Особенностью клеток животного типа (КЖТ) является то, что в условиях культуры ткани, то есть изолирования КЖТ от условий организма, проявляется феномен лимита Хейфлика. Это является одним из вариантов особенностей таких клеток в условиях «вырывания» их из среды сбалансированных факторов, идущих с разных этажей, определяющих статус ювентальности или сенесцента, то есть из общего контекста. Ювентальность или сенесцент регулируется у них через транс- и контрпозитные механизмы иммунных клеток. В условиях изолирования из этого контура регулировок, клетки теряют механизм коррекции со стороны противоположного крыла маятниковой регулировки, это стрессирует клетки, что склоняет маятник в одну сторону. У клеток животного типа имеется особенность - наличие сенесцентных или ювентальных устойчивых фенотипов и их модификатов в пределах конкретных морфотипов. Такого нет у растений, так как этот новый механизм регулирования присущ только клеткам животных. Клетки растений могут легко трансформироваться гормонами, а промежуточный механизм регулировки через закреплённый сенесцент отсутствует. Статуса сенесцента у растений нет. Но есть статус ювенильных клеток, который регулируется только гормонами и некоторыми физическими факторами (свет…). Поэтому у животных сенесцент может корректировать онтогенез, тогда как у растений делают это гормоны. Например, первые этапы сенесцента (в детстве) в иммунной системе (через тимус) могут запускаться кейлонно, исходя из внутренних программ развития данного конкретного типа клеток. Внешний фон созревания организма, фактор целостности (его онтогенеза и прохождения по его этапам) имеет у клеток животного типа приоритетное значение для становления статуса сенесцентных или ювентальных клеток, которых нет у клеток растительного типа. Нарушение механизмов сенесцента может стать причиной клеточной неполноценности, то есть не полной автономности и ускоренной клеточной прогерии. Доказано что прогерия связана с клеточной недостаточностью в её системе самоочисток, то есть нарушен механизм аутофагии, что ведёт к накоплению дефектных белков (прогерина), а это соответствует сенесценту. Клетки попросту не могут переходить по ступеням фенотипии и ориентироваться по внешним факторам регулировок: вырваны из контекста. В условиях организмов животных изменены приоритеты важности сигналов регулировок идущих изнутри клеток или из вне. От этих сигналов зависит быть клетке ювентальной или сенесцентной. В растительном мире эти приоритеты даны клеточному уровню, что облегчает клоновое безлимитное существование даже в культуре клеток. У растений попросту не бывает фенотипа сенесцента, да ещё закрепляющийся на уровне эпигенома. Старение растений инициируется не с клеточного уровня, а общеорганизменного. При этом особенностью выращивания клеток животных в культуре ткани является быстрый «перескок» через все онтостадии и остановка на крайней онтофизисной стадии. — Термин предложил Г. А. Гарбузов.
- Онтодианная линия жизни — онтогенез состоит из последовательных этапов развития, причем длина этих этапов строго определена для каждого вида, суммирование всех этих этапов определяет свою онтодианную линию жизни для каждого вида. — Термин предложил Г. А. Гарбузов.
- Реювентализация — возврат к полной ювенильности клеток, полное восстановление тоти- и плюрипотентности.
- Сенолитики — вещества и препараты, которые способны выводить из строя сенесцентные клетки и способствовать становлению ювентальных.
- УКАМ — Универсальный Клеточный Адаптационный Модус – пролог для раковых опухолей. – Термин предложил Г.А. Гарбузов.
- УКОМ — Универсальный Клеточный Онкологический Модус – пролог для раковых опухолей. – Термин предложил Г.А. Гарбузов.
- Трансанизостаз — это выход функции клетки за пределы репликативного гомеостаза и вход в анизостаз сенесцентной направленности. — Термин предложил Г.А Гарбузов.
- Феноптоз — программа ведущая организм к сенилиту, а затем неизбежного отмирания через анизомалии.
- Эпистресс — эпигеномный стресс, ведущий к смещению паттернов на гистоновой рубашке и нарушающих работу генома.
- Энтропостаз — область выхода систем авторегулировки клетки за пределы клеточного гомеостаза — Термин предложил Г. А. Гарбузов.
- SASP — стареющие клетки выделяют множество факторов, которые называют «секреторным фенотипом, ассоциированным со старением»; является антиподом UASP.
- UASP — это так называемый «секреторный фенотип, ассоциированный с ювентальностью» (понятие предложил Гарбузов Г. А.). Ювентальные клетки выделяют множество специфичных факторов ювентально-ассоциированного спектра секретома, на которых держится гомеостаз фенотипа ювентальности. Альтернативное состояние модификатов (фенотипов), когда вырабатывается SASP, то есть сенесцентно-ассоциированный спектр секретома.
Литература
- Г.А. Гарбузов. Двустороннее взаимовлияние механизмов возрастных сдвигов на уровнях гомеостазов и гормезиса на сенесцент и сенилит. 2025 г. сайт garbuzov.org.
- Г.А. Гарбузов. Взлом иммунных механизмов старения – путь к юности и долголетию. Книга. 2025 г. сайт garbuzov.org.
- Г.А. Гарбузов. Самоуничтожение раковых опухолей. Феномен апоптоза. Книга.
- Г.А. Гарбузов. Эпифиз верховный драйвер онтогенеза: ключ к молодости и старению. Книга.
Книги автора
 ВЗЛОМ ИММУННЫХ МЕХАНИЗМОВ СТАРЕНИЯ — ПУТЬ К ЮНОСТИ И ЗДОРОВЬЮ
ВЗЛОМ ИММУННЫХ МЕХАНИЗМОВ СТАРЕНИЯ — ПУТЬ К ЮНОСТИ И ЗДОРОВЬЮ
Книга знакомит с принципами и методиками противодействия старению. Автор впервые разработал интегральную теорию старения организма, которая рассматривает его в виде взаимодействия двухуровневого процесса как на уровне клеток (сенесцент), так и на уровне всего организма (сенилит). Ознакомившись с материалами, приведёнными в данной книге, вы поймёте, что надо делать, чтобы как можно дольше сохранять молодость, здоровье и бодрость духа.
 ЭПИФИЗ — ВЕРХОВНЫЙ ДРАЙВЕР ОНТОГЕНЕЗА. КЛЮЧ К МОЛОДОСТИ И СТАРЕНИЮ
ЭПИФИЗ — ВЕРХОВНЫЙ ДРАЙВЕР ОНТОГЕНЕЗА. КЛЮЧ К МОЛОДОСТИ И СТАРЕНИЮ
Старение — это многоуровневый процесс как на верхних этажах регулировок (системном), так и на нижнем клеточном. Чтобы адресно разрабатывать методы противодействия и сдерживания общего потока старения (геронтос), необходимо найти правильный ответ — откуда исходят именно истинные первичные начала этого общего процесса. Автор провел глубокий анализ природы старения как в растительном мире, так и в животном. Показано, что этот процесс является двусторонним: снизу он исходит с уровня клеток (сенесцент), причиной является эпигенетическая коммитация на основе онтофизиса; а сверху верховным драйвером этого процесса является эпифиз, а не гипоталамус, как это считалось ранее. Исходя из этих фундаментальных разработок, предложен комплекс методик воздействия на все уровни сдерживания старения. Показана динамика взаимовлияния друг на друга верхнего и нижнего этажей старения. Именно верхний этаж определяет весь спектр возрастзависимых заболеваний (анизомалий). Если мы научимся сохранять «эпифизарную молодость» = неотению, то отодвинем старость.
 ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ. КАК МАГНИЙ И ОКСИД АЗОТА ПРОТИВОСТОЯТ СТАРЕНИЮ
ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ. КАК МАГНИЙ И ОКСИД АЗОТА ПРОТИВОСТОЯТ СТАРЕНИЮ
Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента (на уровне клеток) и сенилита (на системном уровне). Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенилита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые на самом деле не болезни, а естественные анизомалии, то есть механизмы реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), чему посвящена следующая книга автора «Биология молодости».
 САМОУНИЧТОЖЕНИЕ РАКОВЫХ ОПУХОЛЕЙ. ФЕНОМЕН АПОПТОЗА
САМОУНИЧТОЖЕНИЕ РАКОВЫХ ОПУХОЛЕЙ. ФЕНОМЕН АПОПТОЗА
Онкология. Можно ли её вылечить безвредными способами? Автор впервые показывает, что можно и нужно! Для этого нужно запустить механизмы избирательного самоуничтожения раковых клеток в организме на основе клеточных программ апоптоза и аутофагии, которые нужно адресно открыть. Предложена Комплексная Программа из противоположно направленных методов, вычлененных в две фазы и направленных на расшатывание и перебалансировку основных жизнеобеспечивающих механизмов клеток, в т. ч. антиоксидантной « прооксидантной, провоспалительной « антивоспалительной и др. У онкоклеток коридор возможностей самозащиты, их гормезис намного уже, чем у нормальных клеток, у которых Жизненная Сила, Витаукт, намного выше. Такая разность возможностей позволяет создавать особые условия, когда становится возможным запуск механизмов самовыбраковки онкоклеток.
 БИОЛОГИЯ МОЛОДОСТИ, 2020 год. Неоконченная рукопись
БИОЛОГИЯ МОЛОДОСТИ, 2020 год. Неоконченная рукопись
Ознакомиться с ней можно на сайте garbuzov.org.
Все мы хотим жить намного больше, чем имеем возможность, но при этом в молодости, а не бόльшую часть жизни в болезнях и немощной старости. Какова природа стадий онтогенеза и, в частности, молодости и можно ли её продлевать за счёт укорочения старости? Действительно, природа показывает возможности неотении, то есть растянутой молодости. Может ли человек воспользоваться этими же принципами и механизмами? Оказывается, да. Но здесь есть глубокие законы природы, которые надо понимать и управлять ими.
Среди важнейших понятий, определяющих онтогенез, автор впервые показал, что он зиждется на принципах онтофизиса и онтостадийности, которые и являются движущими силами. Сделан глубокий экскурс во всю биологию развития живых организмов, чтобы «раскопать» принципы, по которым можно строить методики сохранения молодости.
Чтобы получить ответы на главные вопросы, необходимо было определить, откуда идут первичные, истинные начала переходов на новые стадии и старение. Существовала путаница, с каких этажей организма исходят истоки старения и почему стадия молодости столь коротка. Чтобы ответить на эти вопросы, пришлось создать концепцию матричных осей, на которых происходит реализация развития организма.
Показано, что существует два направления развития старения: нижний — сенесцент на уровне клеток и тканей и верхний — на уровне общеорганизменных нейрогормональных регулировок — сенилит, которые имеют разные механизмы, но являются двумя рычагами единой матричной системы. Эти потоки сливаются в единый геронтос, который проявляется внешне как симптомокомплекс болезней старости.
На основе этих теоретических посылов автором предложены новые принципы запуска матриц развития на неотению, то есть продления молодости, а значит, и на преодоление болезней старости.
Книгу или Программу-консультацию по заболеваниям Вы можете приобрести по нашему адресу.
Вы можете обратиться с вопросами или за консультацией к Гарбузову Геннадию Алексеевичу на сайте garbuzov.org, по адресу: vitauct@yandex.ru или по телефону 8 (928) 239-13-64.

Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне