


Средний срок доставки от 2 до 6 дней
Книга "Здоровье, молодость, долголетие. Как Магний и Оксид азота противостоят старению"
АННОТАЦИЯ
Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента (на уровне клеток) и сенелита (на системном уровне). Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенелита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые на самом деле не болезни, а естественные анизомалии, то есть механизмы реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), чему посвящена следующая книга автора «Биология молодости».
От автора
Важнейшие регуляторные механизмы основных хронических заболеваний, старения организма и плохой регенерации раскручиваются вокруг дефицита магния и оксида азота. Оказывается, этот повсеместный «дефицит» в возрастзависимых заболеваниях не является дефицитом в общепринятом понимании: его можно считать следствием возрастного снижения чувствительности клеток к ним как на уровне центральных регуляторных систем, так и на уровне тканей, и это своего рода центральная ось, вокруг которой раскручиваются упомянутые проблемы. С помощью супрафизиологических доз можно корректировать дефицит магния и оксида азота, восстанавливая Витаукт — главную Жизненную Силу организма человека. В книге приведены многочисленные примеры и результаты клинических исследований эффективности такой корректировки при широком спектре возрастзависимых заболеваний.
На основе серьёзной аналитики разработана новая концепция механизмов становления этапов онтогенеза и старения организма, которые происходят на двух уровнях одновременно: поток процессов, исходящий с уровня клеток (сенесцент) и поток процессов, исходящий с уровня целостного организма (сенелит). Эти два вектора сливаются в единый процесс старения. Первичные механизмы старения заложены в проявлении возрастного эффекта соматического доминирования, основанного на возрастном генокоммитированном ограничении потенциала стволовых клеток.
Эволюция привела современных животных к лимитированию размеров и роста, а с ними появился и лимитированный онтогенез. Эту ограниченность роста можно обозначить как гомойотопия (от слова «гомойо» — способность всех особей вида или органов в них достигать жёстко заданных величин, например размеров, а «топо» — от слова «топология» — размеры, пространство); а неограниченный тип роста — пойкилотопия (от слова «пойкило» — различный, меняющийся в зависимости от внешних факторов). Все древние животные были пойкилотопными. Именно гомойотопизм представляет собой эволюционное достижение природы по коммитированию как размеров особей, так и числу лет продолжительности жизни и лимиту Хейфлика. Это и определило особенности старения современных животных, механизмы реализации которых заложены изнутри, в клетках, а не зависят от внешних факторов. Одновременно появился механизм сенесцента клеток. До этого его не было в природе. В растительном мире такого механизма сенесцента нет. Топологические видовые параметры, а также онтогенез здесь регулируются на уровне всего организма гормонами — институировано. Элевационные (верхние) механизмы онтогенеза и сенелита институированы, в частности, по оси эпифиз — гипоталамус — гипофиз. Но даже их деятельность обеспечена механизмами коммитации на уровне их клеток. Можно утверждать, что элевационный механизм отключения менструальных циклов (климакс) и сенелита связаны не с лимитом Хейфлика (репликационный сенесцент клеток), а с проявлением коммитирования функции вышеупомянутой оси.
Это означает, что существует отдельно верхний механизм регулировки онтогенеза и времени наступления сенелита. Коммитирование начинается с эпифиза, где уровень выработки мелатонина снижается в 10 раз. Это ведёт к перестройкам в гипоталамусе, гипофизе и железах мишенях. Коммитирование строго определяет наступление менархе (включение цикла) и менопаузу (выключение). Коммитирование определяет и сенелит. Время наступления сенелита далеко опережает время наступления сенесцента, то есть старения, которое идёт снизу с клеточного уровня навстречу сенелиту.
Многие анизомалии (возрастзависимые «болезни») связаны с сенелитом, а часть — со сенесцентом, причём первыми проявляются симптомы анизомалий, связанных с сенелитом, а точнее — с матурой (перезрелости), и намного позже — с сенесцентом.
Матура дословно означает «перезревание», когда гормональный фон за пределами гормезиса, жизненный ресурс резко ослабляется и легко может быть выведен из колеи при различных нагрузках.
К проявлению сенесцента можно отнести сарко-, эндотелий- и эпителийпению и соответственно множество «болезней», связанных с ними.
Очевидно, что старение, или сенелит, имеет три источника, направления: это сенесцент, коммитация и элевационное институирование. В конечном итоге все эти три направления с разными механизмами сливаются в единый поток. Следовательно, неверно будет вести поиск какого-то специфического средства; решение должно быть многоплановым и всесторонним (плейотропным).
В то же время надо вести фронт по следующим направлениям:
— во-первых, с сенесцентом, например, с помощью мегаантиоксидантов и инфламантов (противовоспалительное направление);
— во-вторых, надо противодействовать процессам, которые начинают раскручиваться на уровне эпифиза, восстанавливая уровень мелатонина и синтропийных с ним нейрогормонов;
— в-третьих, вести разработки по противодействию коммитирования, то есть путём перепрограммирования стволовых клеток.
Чтобы прийти к таким выводам, автор попытался выстроить свою интегральную теорию биологии старения, объединяющую самые различные разрозненные научные данные о старении в мире живого — как в царстве растений, так и в животном царстве, а также показать многогранность и многоуровневость явления в пределах организма. Это позволило устранить противоречивость ряда концепций и уложить ряд пазлов в единой картине науки «биология старения».
Также впервые дано новое понимание сопряжённых с возрастом «болезней» старости, которые с научных позиций эволюции онтогенеза не являются таковыми, и их правильнее определять как анизомалии, то есть выход за границы оптимума для клеток и систем, называемого также коридором гормезиса — своеобразного «русла», в котором все процессы сохранения здоровья обеспечиваются силами Витаукта (надо понимать, что здоровье не может поддерживаться за пределами данного коридора). Возрастная нечувствительность клеток связана со смещением их констант гомеостазов. В результате сигналинговая система клеток нарушается, и в первую очередь это сказывается на чувствительности клеток к регулирующему действию основных клеточных сигналинговых веществ — магния и оксида азота. Только в пределах «русла» гормезиса, интегрирующего все потоки жизнедеятельности различных систем и обеспечивающего Витаукт, а также сохранения констант гомеостазов клеточного уровня, обеспечивающих Жизненную Силу на уровне тканей и органов, возможно поддержание в оптимуме функциональности на всех «этажах» такой сложной биологической системы, как организм человека.
Неизбежность анизомалий возможна только в мире животных. Они являются реализацией внутренних программ онтогенеза и инструментами феноптоза — мортального исхода. Анизомалии никогда не поддадутся простым медицинским терапевтическим методам и коррекции, поскольку последние не затрагивают истинные генные первоначала. Анизомалии во многом сходны с рядом хронических заболеваний, которые тоже могут вести к преждевременному старению, так как имеют единые вторичные механизмы реализации. Обычные болезни (кроме генетически врождённых) преимущественно имеют экзогенные начала. Причём хронические их формы работают через те же механизмы воспаления и другие механизмы, так же, как и при сенесценте клеток влияют на эпигеном, укорочение длины теломер и митохондриальные дисфункции, истощают эрготропный ресурс, тем самым дополнительно вмешиваются, вливаются в общие первичные запрограммированные процессы развития и обеспечивают преждевременное старение на клеточном уровне.
Часть анизомалий связана с сенесцентом, а часть — с сенелитом.
Клеточное старение определяет старение тканей, органов и организма. В частности, по этой причине происходит искажение функционала высших регулировочных систем с выходом за пределы оптимума.
В свою очередь старение организма как системы систем (нейроэндокринный уровень) дополнительно накладывает свой отпечаток на темпы эндогенного старения клеток. Эндогенное старение клеток сцеплено как с лимитом Хейфлика, так и с клеточным соматическим доминированием над стволовыми клетками и чрезмерным коммитированием прогениторных клеток. Одно определяет другое по принципу замкнутых обратных связей. Особенностью старения организмов высших животных, в отличие от растительного мира, является блокировка универсальных программ различной степени потентности, включая тотипотентную к регенерации и репарации, а также ограниченные возможности прогениторных стволовых клеток.
Теория определяет практику. Автор считает, что в случае, если удастся создать правильное понимание этой проблемы, то только тогда возможно начать поиск в нужном направлении и выйти на правильный путь решения задачи продления жизни человека. По мнению автора, этот путь заключается не в сдерживании старения и противодействии ему (это возможно только для преждевременного старения, связанного с болезнями, но не с анизомалиями), а в подключении механизмов неотении — продления молодости. Эта книга является прологом к следующей книге автора, которая будет называться «Биология молодости».
Центральная роль магния в клеточных гомеостазах организма
Анализы часто могут показывать норму Mg в плазме, но при этом его может существенно не хватать в костной и других тканях, функциональность которых сильно связана с Mg. Здесь можно говорить о скрытом дефиците Mg, причины которого нам малоизвестны.
Среди всех катионов магний занимает 4-е место по содержанию в организме и 2-е место по содержанию в клетке. До 53 % магния концентрируется в костной и зубной тканях, около 20 % — в тканях с наиболее высокой метаболической активностью (мозг, сердце, мышцы, почки, печень).
Магний — важнейший макроэлемент, и это связано с тем, что, в отличие от других элементов, таких как калий, натрий, кальций, он не просто строительный материал, а основная регуляторная ось клеточных гомеостазов, относительно которого балансируются = подстраиваются все остальные. Каждый гомеостаз регулируется двумя противоположными рычагами. Такими противоположностями являются Mg « Ca. Это дуэт антагонистов. Это единство и борьба противоположностей. Ни один из них не может полноценно работать в отсутствии или недостатке противоположного. Каждый из них имеет своё полезное предназначение в пределах единой маятниковой системы, где ось этого маятника представляет собой константу гомеостаза. Каждый рычаг маятника представляет собой транспозитную силу и контрпозитную силу, тем самым очерчивая собой круг возможностей выполнения определенных функций. Все параметры их величин, то есть оптимизированные константы определены своими программами. Это некое поле, зона, которым они управляют или реализуют. За пределами этой зоны гомеостат не работает.
Каждый гомеостаз имеет сопряжение с другими гомеостазами через триггерные гены. Все они интегрированы в единую систему гомеостазов и имеют взаимооптимизированные константы.
До сих пор при возрастном остеопорозе, то есть недостатке кальция в костях, предлагают применять кальциевые препараты, что, исходя из правила полярности гомеостаза, в принципе неверно и способно, кроме небольшой пользы, принести намного больший вред в виде кальцинозов.
Относительно нормо-показателей Mg (константа гомеостаза) ориентированы константы всех остальных минералов. Нет отдельных независимых гомеостазов по Na, Ca, К и т. д., а есть единый интегрированный водно-минеральный гомеостаз с множеством параметров, в центре которого находится константа Mg, относительно которой как в вихре подстраиваются, регулируются с помощью различных систем другие константы. Если сбита основная константа, то происходят компенсаторные процессы подстроек в других параметрах. Если константа Mg в норме, то под неё ориентируются все остальные, а не наоборот.
Относительно уровня Mg зависит или регулируется с его помощью энергетика клетки. 90 % магния задействовано в клетке на образование и поддержание АТФ в митохондриях — энергетической валюты клетки. Становится понятной его центральная роль в энергетической системе клетки, от мощности которой зависят все остальные функциональные возможности клетки. Участвуя в работе сотен ферментов, он определяет направленность синтеза множества белков, жиров и углеводов.
Почему с возрастом клетки стареют и становятся невосприимчивыми к обеспечительным субстратам?
С возрастом происходит неотвратимое снижение уровня содержания магния во многих клеточных депо. Особенно в костях в виде неизбежного старческого остеопороза. Почему? А ведь Mg является одним из ведущих ростовых факторов и оптимизаторов существования клеток. Чтобы дать ответ на природу этого неизбежного процесса, необходимо дать ответ, откуда исходят его начала. Первое возможное направление их истока можно связать с гормональными возрастными перестройками, когда процесс спускается сверху вниз. Но есть множество фактов, идущих вразрез с этим утверждением и показывающих, что все его истинные первопричины следует искать в старении на клеточном уровне… Как итог — их дисфункция, снижение жизнеспособности и чувствительности к регулировочным сигналам, резистентность, а точнее ослабление ВИТАУКТ = Жизненной Силы. Затем к этой резистентности подстраиваются верхние уровни нейрогормональных регулировок в виде элевационных подстроек, то есть наращивание мощи гормональных сигналов с целью преодолеть барьер нечувствительности к ним. Можно утверждать и еще точнее, что первые этапы онтогенеза организма (постнатальный, ювенильный…) движутся с помощью гормональных перестроек, то есть исходят с верхних этажей иерархии регулировок, а на последних этапах — как адальтус (зрелость), матура (перезрелость), сенелит (старение) — подключаются и иные процессы (сенесцент — клеточное старение), которые исходят из клеточного уровня, обусловлены порогом Хейфлика, ведущего к исчерпанию их возможностей и податливости к регулировкам, то есть появления резистентности на их уровне, что и лимитирует их функциональные возможности.
С возрастом неотвратимо накапливается количество сенесцентных клеток, которые обеспечивают эффект соматического доминирования вместо в норме прогениторного доминирования, то есть одновременной работы в разных тканях множества типов различных стволовых клеток-инициаторов на непрерывное самообновление ткани в целом. В связи с новым пониманием проблемы возник новый альтернативный подход, показывающий важность сенолитиков для сенолитического преодоления старения на клеточном уровне.
В экспериментах на культуре ткани клеток показано, что недостаток Mg ведёт к ускорению старения клеток. Но можно предполагать и противоположное, то есть утверждать, что клеточное старение (сенесцент) приведёт к последующему снижению восприимчивости к Mg. Может быть, ответ в том, что линии прогениторных клеток животных даже в идеальных условиях имеют свой предел (предел Хейфлика) и это неизбежно включает программы угасания энергетики и выхода на сенесцент. Это не энтропийно-стахостический, то есть процесс случайностей и изнашивания, а реализация, исчерпание ограниченных возможностей, заложенных в генных программах противодействию как внешней энтропии, так и внутренним процессам возрастного самоизживания. Это клеточная инволюция. Инволюция начинается не на уровне организма, сверху на уровне гормонального баланса, как это считалось раньше, а на клеточных линиях стволовых клеток, что в свою очередь приводит к нарушениям и на верхних регулировочных уровнях. Идёт тотальное, повсеместное угасание стволовых клеток во всех системах и тканях.
Что означает термин «самоизживание»? На уровне организма процесс имеет своё понимание, а на уровне клеток — другое. На уровне организма это процесс перехода зрелости в перезревание организма (матуры), когда запускаются механизмы климактерия, а затем сенелита. Всё это завязано не с лимитом Хейфлика, а с инволюционным эффектом Хейфлика, который, в свою очередь, синтропиен системным элевационным процессом в организме. Синтропия — это обязательно сопряжённые или идущие параллельно процессы.
Одно обуславливает другое, одно предназначено для реализации другого. Нижний клеточный уровень определяет возможности верхних этажей, все зациклены в одном круге. Ослабление потенций в нижней части круга ослабляет функциональные возможности верхних уровней.
В каждом органе и ткани имеются свои независимые мезенхимы и прогениторы, где идёт курс на ослабление роли мезенхимы и усиление прогениторов. Каждая ткань стареет по своим внутренним правилам очерёдности и в тоже время соподчиняется общей тенденции. Этот же процесс происходит и в верхних центрах регулировки. Как итог, смещение гормонального профиля, который выходит за пределы оптимальные для организма в целом. Это в свою очередь отражается на нижние уровни тканей мишеней, в которых начинают закладываться основы и предрасположенность для многочисленных заболеваний возраста и сенелитирования, в итоге переводит организм из состояния матуры в сенелит. Здоровая старость в принципе невозможна.
В наших организмах не предусмотрены механизмы противодействующие сенелиту путём непрерывной, беспредельно самоподдерживающейся неотении (самоомолаживания), которые могли бы отодвигать эти процессы. Соотношения сенелита и неотении — это программируемые процессы, приобретённые в ходе филогенеза в нашей эволюции. Лимит Хейфлика обуславливает сенесцент, а элевационный лимит определяет сенелит. Следует отметить, что численное количество потенциала циклов митозов приблизительно схожее как у животных длительно живущих, так и у короткоживущих. Сенесцент ограничивает клеточную Жизненную Силу, а сенелит ограничивает Витаукт; сенесцент смещает точки координат = константы гомеостазов, а сенелит смещает гормезис. С сенесцентом связано нарастание возрастной клеточной рефрактерности, вторичной нечувствительности, в частности к Mg и недовыработки NO в митохондриях. Похоже с этим связана элевационная нечувствительность гипоталамо-гипофизарной регуляторной системы, контролирующей и сохраняющей гормезис. Но эта система находится в непрерывной динамике, обеспечивающей элевационные механизмы, которыми движут как рост обратных гормональных ответов, то есть экзогенных, так и рост сенесцента в этих же регуляторных структурах, то есть эндогенных. Рост обратных гормональных ответов имеет свой допустимый порог для их уровня в центральных регуляторных структурах, затем начинается подстройка под ситуацию высших регуляторов — это и есть элевация. Происходит непрерывное противостояние транспозитных сигналов снизу и контрпозитных сверху. Элевация в лимитированной по росту системе приводит к серьёзным качественным смещениям статуса организма, утрате устойчивости и выводу его функций за пределы оптимума.
Иначе говоря, это означает переход адальтуса (зрелость) в матуру (перезрелость), а затем в супраматуру (супраперезрелость с переходом в сенелит) или выход за пределы оптимума, то есть за коридор гормезиса, когда набирает силу всё множество возрастных болезней. Всё это обусловлено ограниченными возможностями лимитированных форм. Здесь длительность жизни варьирует между возможностями продления неотении и неизбежной потребностью входа в фазу генеративной = половой зрелости, которая в свою очередь дополнительно ограничивает прогениторную силу стволовых клеток. Замкнутые ювенильно-ювенальные системы в принципе устойчивы, но нерациональны эволюционно, так как должны себя поддерживать расширением роста наружу, например вегетировать, либо постоянно производить внутри себя самообновление, самоомолаживаться, при этом мощно тормозить процессы роста. Два взаимоисключающих процесса. При непрерывном росте нужны механизмы репликации, а при самоомоложении — репарации. Репарация может происходить путём аутофагии старых клеток и замены на новые либо репараций внутри ядра или сомы клетки. Природа пошла по пути компромисса, то есть удерживать свою устойчивость как репарациями, так и управляемым процессом непрерывного повышения качественных изменений за счёт повышения степени половой зрелости, то есть отмены сохранения уравновешенного состояния поддержки заданного уровня зрелости. Это неизбежно ведёт к её нарастанию и переходу в адальтус, а затем в матуру и климакс. Этот переход обеспечивает нарастание нечувствительности = рефрактерности в центральных звеньях регулировки. Зрелость — это компенсация закрытого неограниченного роста, которая направлена не во вне, а внутрь. Ведущими здесь являются центральные структуры регулировки, обеспечивающие элевационные процессы.
Особенностью растений является возможность сохранения одновременно возрастной разнокачественности побегов и почек (ростовых точек), например, на разных частях дерева имеются и ювенильные (тотипотентные) точки роста, и зрелые (проявляющие унипотентность, например, в виде топофизиса или циклофизиса). Подробности в книге Г.А. Гарбузова [2]. Такая организация организма даёт растениям ряд преимуществ, например, возможность неограниченно регенерировать и создавать новые побеги в любых местах организма или самоомолаживаться, то есть полностью воссоздавать ювенильные ростки аналогичные тем, что происходят из семян.
В свою очередь у животных такая онто-возрастная разнокачественность закрыта полностью, она ограничена на уровне всего множества типов прогениторных стволовых клеток. С этим связано отключение возможности как к органной регенерации, так и пролонгированной неотенизации у высших животных. Эта же закрытая прогениторность обуславливает как сенесцент, так и сенелит. Организм животного — замкнутая система, где невозможно оптимально управлять вне рамок возможностей прогениторности. Организм развивается непрерывно внутрь самого себя, а не вширь, чем и изживает самого ебя.
Особенность растений в том, что их вегетативные потомства могут иметь неограниченные возможности к делениям и существованию, у них нет программ сенесцента, то есть сугубо на клеточном уровне, но таковая обнаружена в клонах клеток животных. Возможности генетических программ растений определены тем, что они фактически являются открытыми системами и способны перезакладывать в новом месте как свои апексы — меристемные верхушки точек роста, так и неограниченное число спящих почек (ювенильных). Таким образом, растения — представители открытых систем, то есть с потенцией к неограниченному росту. Такой потенции неограниченного простого роста у прогениторных клеток животных нет. Это определено в филогенезе, и ограничения связаны с тем, что их стволовые клетки способны к восстановлению новых потомств, но при условии, что родительские стволовые клетки должны уйти, освободить место потомкам или быть постоянно подавленными и работать в реципрокном режиме, то есть быть в подчинении у соматических клеток. Реципрокность регулируется как верховными сигналами, так и исходящими изнутри клеток. Каждый новый реципрокторный цикл должен запустить программу заново и синтропиен с указаниями из соматической клетки. Таким образом, здесь главенствуют соматические клетки, то есть принцип соматического доминирования. Всё подстроено под обеспечение их непрерывного существования, вплоть до пожизненного существования, например у нейронов. Организм животного обязан поддерживать постоянство своих форм и размеров. Для их сохранения организм животного вынужден жертвовать количественными переходами на качественные, что можно обеспечить за счёт цепи переходов с одного этапа на другой, когда неизбежно старение.
Например, в костной ткани всегда сохраняется баланс между остеокластами (клетки, занимающиеся в основном резорбцией костной ткани) и остеобластами (клетки, обеспечивающие формирование костной ткани и минерализацию).
За счёт этого происходит обновление клеточных потомств, но при этом сохраняется размер и форма костной ткани. Но такое обновление потомства клеток существенно отличается от подобного у многолетних растений. Функции резорбции или замкнутой реципрокности для них не нужны, новые клетки переоткладываются на новом месте, не разрушая старые. Потенциально такое растение может расти неограниченно, например с помощью вегетативного или клонового размножения. Здесь работают открытые реципрокные (самовозвращающиеся) механизмы возврата к прежним программам циклического роста. Открытая реципрокность подразумевает принцип любого типа апикального доминирования. Эти циклы, если их не ограничивать другими программами, могут быть бесконечными. Стволовые клетки переоткладывают сами себя в каждом цикле.
У животных клеток реципрокность регулируется преимущественно с внутриклеточного уровня; это не та открытая для внешних стимулов реципрокность, как у растений, которая позволяет им открыть полностью все степени потентности для регенерации. У животных эта потентность глубоко подавлена и проявляется на уровне прогениторных стволовых клеток. Возврат в проэмбриональное дедифференцированное состояние им не дано, следовательно, нет и полного цикла самоомоложения клеток в реципрокторных циклах. В этих циклах сохраняется большая часть степени их дифференциации. Невозможность освободиться от груза дифференциации приводит к созданию «приложений» программ, то есть происходит запуск специальных дополнительных программ для автостартирования следующего цикла.
Итак, у животных реципрокность регулируется не с уровня потребностей и потенций верхних этажей гормонального регулирования (институативно), а изначально изнутри на уровне определяющих их потребности соматических клеток (коммитированно), которое реализуется за счёт механизмов, предначертанных для каждого типа стволовых клеток (конституативных).
Возможен и другой принцип, когда у клеток животного типа весь геном должен быть закрыт, а клетки-носители её уничтожены, но затем открывается заново в новом потомстве из обычных клеток, когда они могут вернуть себе способности им предшествующих прогениторов. Это так называемый реверс, когда соматические переходят в прогениторные, а последние создают соматические. В целом процесс протекает реверсно, то есть с помощью других аналогичных программ. Каждый раз клетки не копируют себя с сохранением той же самой программы, а включают заново версию той же программы. Родительская версия уходит с родителями, а в потомстве перезапускается её трансляция. Запускают её уже другие механизмы через гены триггеры. В каждом очередном митозе идёт триггерный перезапуск. Возможно, прежний триггер недоступен, он заблокирован программой соматической дифференциации. Каждый новый цикл едет как по «шестерёнке» и зацепляет новый её «зуб» = триггер.
В существующей концепции лимита Хейфлика есть свои «дыры», которые ею не могут быть объяснимы. В частности, у растений нет такой клеточной потенции на сенесцент и не срабатывает этот механизм. Почему у них нет этого счётчика? Почему фермент теломераза, отвечающий за восстановление теломеры, может восстановиться и отодвинуть порог Хейфлика? Почему мыши с многократно раз более длинной теломерой и не заблокированной теломеразой способны на меньшее количество митозов, а продолжительность их жизни многократно раз меньше?
Для возобновления клонового потомства клеток животных не используются универсальные тотипотентные программы (эмбриональные), а опосредуются через вторичные плюри- и мультипотентные программы (взрослые стволовые клетки). Это частные программы, работающие только на данную фенотипическую популяцию клеток, отвечающих за поддержание именно данных, конкретных морфогенетических линий клеток. Запуск мультипотентных программ возможен только через триггерные программы — при условии, что родительские клетки освободили место для потомков. То есть открытой цепочки к возможности непрерывного деления здесь нет. Причём для каждого нового цикла возобновления потомства должна быть предписана своя программа заново. В итоге получается, что это совокупность программ, а не одна программа к рециклингу. В филогенезе оптимизирована сумма этих возможных самовоспроизведений. Но она не может, да этого и не нужно, быть неограниченной, то есть открытой. Поэтому, в отличие от растений, способных к неограниченному воспроизводству клонов, у животных это клонирование, количество циклов воспроизведения ограничено, причём регулировка происходит не на верхнем уровне организма, а на изначальном, клеточно-генном. Их возможности предопределены и зафиксированы. Клетки животного типа обязаны существовать только в замкнутых пространствах определённых форм, то есть стволовые клетки должны существовать только в пределах системы закрытой морфогенетической ткани. Возможности регенерации здесь фактически нет, или она сильно ограничена, а только определенная репарация. Это означает, что стволовые прогениторные клетки имеют внутренние генетические самоограничения, у них строго запрещены тотипотентные = универсальные программы, обеспечивающие тотальную регенерацию. Для этого им надо было бы сбрасывать полностью всю надстройку комплекса высших программ и уходить в зиготное состояние или эмбриогенез, что для них исключено.
При этом есть основания утверждать, что в зиготе все программы открыты и ждут своего сигналинга, толчка для последовательного трансформинга; в стволовых клетках всё наоборот — открыты (экспрессированы) только те гены, которые обеспечивают данную высшую функцию клетки, остальные репрессированы.
Каждый новый переход стволовой клетки на такое зиготоподобное тотипотентное состояние должен быть обеспечен полным комплектом программ обеспечения для возврата в исходное состояние и новых программ на дифференциальный рост. В принципе такая клетка в замкнутой системе должна иметь бесконечное число триггерных программ на обратный исход в изначальное состояние и затем возврат. Но природой не предусмотрено иметь столько множество мультипликативных программ. Постепенно запас этот исчерпывается, так как здесь работают только плюрипотентные, мультипотентные и унипотентные программы, но исключены тотипотентные. В пределах замкнутого морфогенеза тотипотентность невозможна. Эта генетическая предопределенность влияет и на особенности фенотипа онтогенеза, индивидуальную среднюю продолжительность жизни данного вида животного и на неминуемые процессы феноптоза (программа на отмирание особи). Одним видам животных предусмотрена продолжительность жизни 1–2 года, а другим — до 100 и более лет. Конечно, эти особенности фенотипа онтогенеза определяются не только на клеточном уровне, но могут определяться и на общеорганизменном уровне, который через ухудшение нейрогуморальных регулировок может определять клеточное ускорение наступления климакса и затем включение мортальных механизмов феноптоза. Например, таковыми являются механизмы умирания рыбы горбуши, когда в девятилетнем возрасте она заходит в пресноводные реки на нерест, и в это время её уничтожают внутренние механизмы остро проявленного быстрого мортального нейрогуморального климакса, которая по сути является стадией матуры онтогенеза, то есть гиперзрелости = перекос гормонального фона, выходящего за рамки гормонального оптимума, то есть гормезиса. Стадии растянутого сенелита = старения здесь практически нет.
Что общего между понятиями онтофизис у растений и соматическое доминирование у животных тканей?
Разнокачественность побегов у растений может проявляться в виде:
— топофизиса — сохранение у клоновых побегов того морфологического строения, которое было присуще ранее побегу в кроне, например плагиотроный (боковой) или ортотропный (прямой) рост;— циклофизиса — способности клоновых побегов сохранять генеративную зрелость в потомстве клонов, например закладывать цветочные почки или стробилы (шишки) у хвойных;
— онтофизиса — способности клоновых побегов сохранять возрастную зрелость или особенности морфологии, присущие для различных этапов существования растения.
На рисунках 1 и 2 показана разнокачественность побегов из различных частей кроны и их клонового потомства из укоренённых черенков. Таким образом, видно, что у вегетативного потомства сохраняются признаки зрелости и морфологии материнской особи.
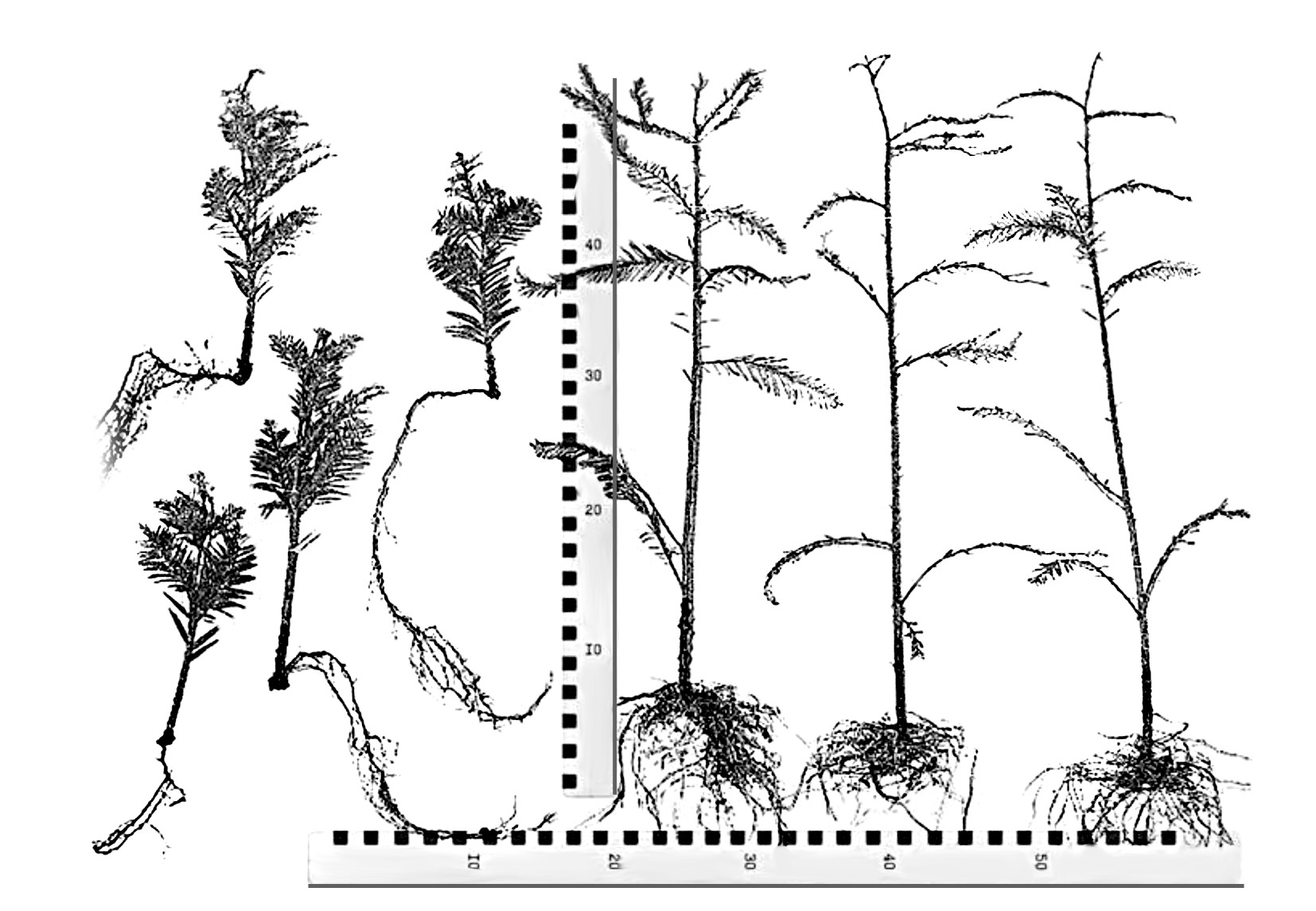
Рис. 1. Справа — внешний вид укоренённых черенков-саженцев от ювенильных ортотропных побегов, заготовленных в ювенильном маточнике, которые сохраняют все признаки ювенильного строения сеянцев и высокой степени к регенерации. Слева — укоренённые черенки от побегов, взятых в кроне.
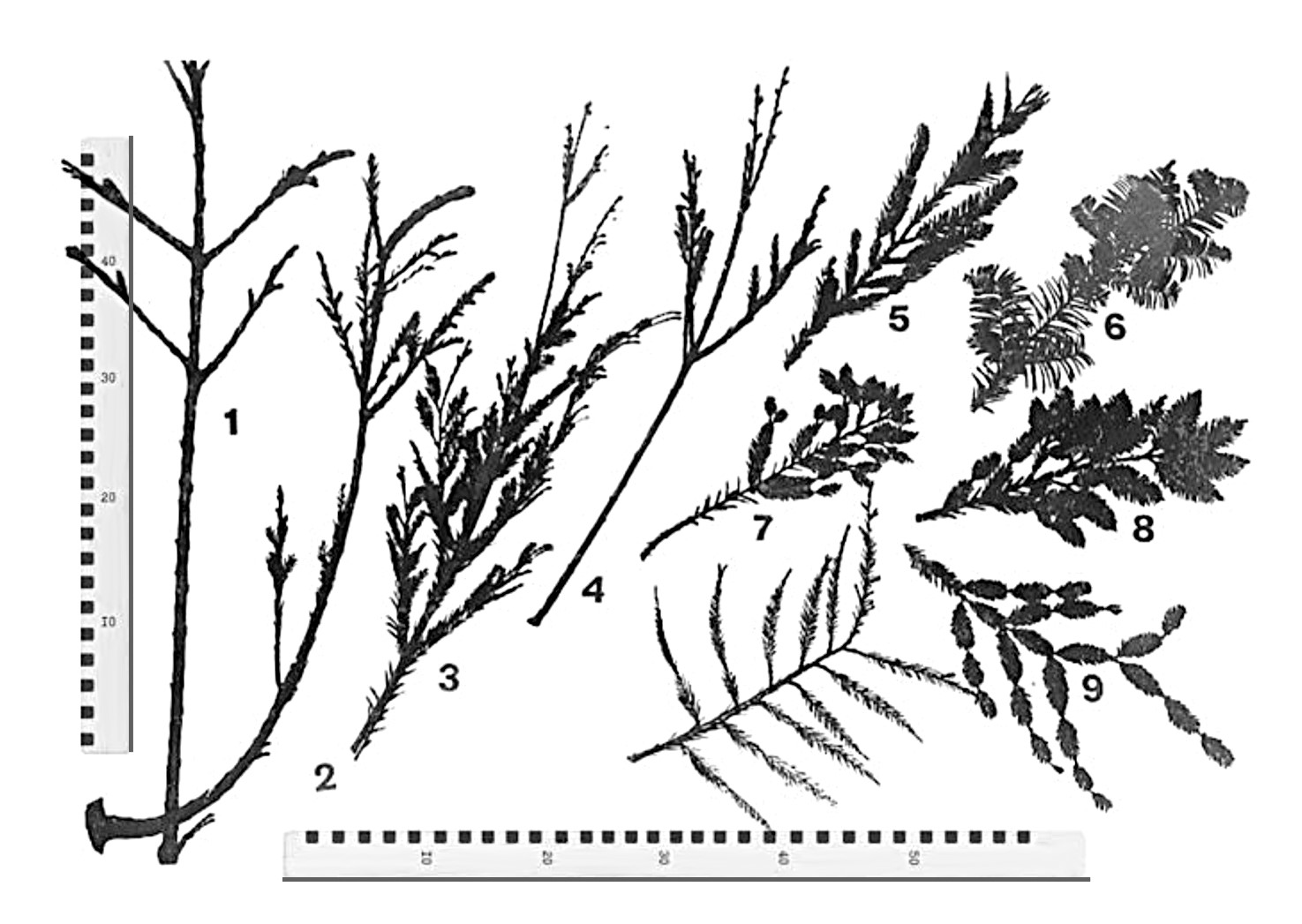
Рис. 2. Широкая морфологическая разнокачественность побегов в зависимости от их расположения в кроне.
У растений все ткани и побеги имеют субординацию и соподчинены апикальной точке роста или главному побегу. Так задаётся морфогенез и зрелость побегов. Все побеги существуют не по своим программам, а скоординированы общим принципом единого целого.
С устранением апикального доминирования, то есть главного побега, например у ели, остальные побеги длительно или навсегда сохраняют статус прежнего соматического состояния в виде экспрессии или репрессии генопрограмм, обеспечивающих статус топофизиса, циклофизиса и онтофизиса. Но, тем не менее, это не является программами сенесцента, то есть старения на изначальном клеточном уровне, то есть на терминальных = концевых стволовых прогениторных клетках, а является следствием созревания организма в целом, когда он переходит с этапа ювенильности и вегетативного роста на следующий этап онтогенеза — адальтус, то есть гормональной зрелости, способной запускать программы на создание генеративных органов. Это означает, что их меристемы в апикальных точках достигают таких достаточно высоких величин, когда способны вырабатывать объём гормонов, способный преодолеть порог нечувствительности к ним в рабочих тканях и достаточный для включения программ на образование генеративных органов и радикальной морфогенетической перестройки. Таким образом, количество переходит в качество, когда становится возможным экспрессировать, открывать новые программы на создание генеративных органов.
У тканей животных всё обстоит несколько иначе: каждая из тканей представляет собой определённую автономию и самоуправляющиеся структуры и в меньшей степени зависит от общеорганизменных регулировок. Животные имеют многоцентровые зачатки ткани, которые развиваются более независимо по своим программам морфогенеза. Тем не менее, у них тоже имеется верховный координатор, который ведёт себя аналогично апикальной меристеме у растений: после ряда становлений в них происходит генеративная зрелость, которая и запускает зрелость или гормональные перестройки на железах и тканях мишеней. В каждом таком фенотипе (с автономными признаками) ткани происходят как свои онтогенетические перестройки = созревание, так и за счёт внешних регулировок. Всё это вместе приводит к некоему подобию онтофизиса внутри клеточных дочерних линий. Основные типы потентности у них практически заблокированы, кроме унипотентности в прогенитоных типах клеток. Таким образом, даже тот небольшой уровень потентности с возрастом у них тоже суживается.
Роль сенесцентных клеток в возрастной рефрактерности клеток
- Доставка почтой России
- Курьерская доставка (до двери)
- Доставка транспортной компанией (СДЭК, Boxberry, ЦАП, Деловые Линии)
- Самовывоз по адресу г.Сочи, ул. Северная 6.
Обратите внимание, что у Почты России есть три тарифа: Обычная доставка, Ускоренная и EMS.
Ускоренная – на 80-100% дороже, в среднем посылка приходит на 2-3 дня быстрее.
EMS – доставка посылки до двери.
Вопросы по доставке вы можете задать по телефонам:
+7 (938) 467 78 06
+7 (995) 003 74 85
Стоимость доставки
При заказе обычной доставки почты России у нас действует система скидок. Чем больше ваш заказ, тем ниже стоимость доставки. Обратите внимание, акция действует только на территории РФ.
Сумма заказа
|
Сумма заказа |
Стоимость доставки |
|
До 800 рублей |
350 |
|
От 800 до 3500 рублей |
300 |
|
От 3500 до 5000 рублей |
250 |
|
От 5000 |
Бесплатная доставка |
При ускоренной доставке Почты России, EMS или транспортной компании, Вы оплачиваете фактическую стоимость доставки. Она зависит от расстояния и размера заказа. Стоимость доставки рассчитает наш сотрудник, который с Вами свяжется.
- Переводом на карту Сбербанка 2202 2085 3388 6520, владелец Геннадий Алексеевич Гарбузов. Платеж можно сделать с помощью приложения Сбербанк Online, через банкомат или в отделении Сбербанка (все варианты без комиссии). Или переводом с карты другого банка (размер комиссии уточняйте в вашем банке)
- Произвести оплату с вашей банковской карты при оформлении заказа (через сервис Robokassa)
- Произвести оплату с вашей банковской карты по выставленному счёту (через сервис Robokassa).
- Оплатить с помощью платёжных систем Золотая Корона (для жителей СНГ), Western Union и MoneyGram (для жителей дальнего зарубежья)
- Возможны иные способы оплаты по согласованию с вашим менеджером.
Обратите внимание! Заказы мы отправляем только по предоплате. Если Вам по какой-то причине это неудобно, мы готовы пойти на компромиссный вариант: Вы внесёте частичную небольшую предоплату, а остальную сумму оплатите при получении посылки.
Возврат или обмен товара, который не подошел (не понравился), допускается, если с момента покупки прошло не более 7 дней (статья 26.1, пункт 4 ЗоЗПП) и он не входит в перечень товаров, не подлежащих возврату и обмену. Для проведения операции нужно позвонить по номеру +7(495)744-63-63 или отправить письмо на электронный адрес vitauct8@yandex.ru.
Обменять или вернуть товар можно в течение недели, если соблюдены условия:
- товар не использовался, не вскрывался;
- упаковка не повреждена;
- не нарушены технические, лечебные, другие свойства товара;
- сохранен товарный вид изделия;
- присутствуют ярлыки, пломбы;
- есть документация на продукцию, чек на покупку.
Обмен производится однократно на аналог такой же стоимости (без перерасчета) или на более дешевый/дорогой аналог с перерасчетом стоимости и возвратом/доплатой.
При отказе от заказа Garbuzov.org обязуется вернуть уплаченную по договору сумму в течение 10 дней со дня предъявления требований по возврату, за исключением расходов на пересылку товара, которые оплачивает покупатель (статья 26.1). При ненадлежащем качестве продукции продавец берет на себя транспортные расходы в оба направления.
При 100 %-ной оплате заказ нужно получить до истечения срока хранения. Если заказчик не забирает посылку в установленные сроки хранения, товар возвращается отправителю. Транспортные расходы в этом случае оплачивает заказчик.
Подробную информацию можно узнать здесь.





Попробовали? Оставьте отзыв