
Биология молодости
КАК ЗАПУСТИТЬ МЕХАНИЗМЫ ОМОЛОЖЕНИЯ = РЕГЕНЕРАЦИИ И РЕЮВЕНИЛИЗАЦИИ = ВИТАЛИЗАЦИИ = УСИЛЕНИЯ ЖИЗНЕННОЙ СИЛЫ = ВИТАУКТ = УСТОЙЧИВОСТИ К БОЛЕЗНЯМ СТАРЕНИЯ
ЭКЗОГЕННЫЕ И ЭНДОГЕННЫЕ МЕХАНИЗМЫ СТАРЕНИЯ И БОЛЕЗНЕЙ СТАРОСТИ.
Что такое симптомокомплекс старения и синдроматика старения и как им противодействовать?
Старение – этой двусторонний процесс, то есть идущий как с уровня генетических врожденных программ и являющийся одной из частью жизненного цикла = онтогенеза, а также как как проявление накопления ряда эпигенетических изменений, приобретенных в течение жизни. Это объясняет почему у старения есть общие универсальные черты, присущие для всех, и индивидуальные особенности, связанные с эпигенетическими болезнями возраста. О старении = сенелите как о части онтогенеза мы будем обсуждать отдельно, но также отдельно рассмотрим ту его часть, которая приобретена при жизни, и укорачивает нам её. Это так называемый Симптомокомплекс Болезней Старости = СБС. В принципе если бы не СБС человеческий организм способен прожить в несколько раз больше, чем средняя продолжительность современных людей = 70 лет. К таким ярко выраженным симптомам старения относятся:
- саркопения – мышечное вырождение, атрофия,
- меланопения - седые волосы = исчезновения стволовых клеток, вырабатывающих краситель меланин,
- нейропения – уменьшение нервных клеток,
- кератопения – проблемы с коллагеном в коже,
- гепатопения – в печени,
- эпителиопения - эпителиальная дисфункция стенок сосудов, ведущая к атеросклерозу, отмирание до 50% микрокапилляров и т.д.
Практически во всех тканях и системах организма идёт относительно быстрое или медленное изменение баланса деятельности стволовых клеток и их угасание, но причина которых на эпигенетическом уровне. Если нам удастся оживить активность этих стволовых клеток, преодолеть эпигенетику, то сможем остановить такое старение и продлить здоровую жизнь вплоть еще на 100 лет.
К синдроматике сенелита можно отнести климакс – неминуемый гормональный сдвиг, а также весь комплекс процессов, связанный с нарушением, угасанием активности стволовых = плюрипотентных клеток.
Верхние и нижние этажи организма, где зарождаются истоки механизмов старения.
Большая часть методик и рекомендаций по сдерживанию старения касаются болезней старости, например атеросклероз, относится к их экзогенным началам, которые в какой-то мере можно корректировать. Если они как, то и действуют на наш геном и его программы, то в основном на эпигенетическом уровне = приобретённом.
Но есть и радикальные причины болезней старости, в том числе и атеросклероз, которые сцеплены генетически с этапами онтогенеза и в большей степени связаны с гормональными перестройками на общеорганизменном уровне, а также энергетическими и ростовыми ограничениями на клеточном уровне. Все они вписаны в генетические программы развития и не поддаются коррекции.
Примером таких изменений на общеорганизменном уровне является уменьшение в 10-ки раз выработки организмом нейромедиатора мелатонина, который присущ для этапа неотении = юности, но не для сенелита = старости. Таким образом, болезни старости осуществляются на фоне существенного изменения гормонального профиля. Рассматривая проблему с этого ракурса, казалось бы, корни ее, первичные механизмы начинаются на верхних этажах организма. Но тогда следует ответить на вопрос что приводит к изменению с возрастом гормонального профиля. Основной механизм регулировки обеспечивает эпифиз. А кто же тогда управляет возрастными изменениями в эпифизе? Легче всего предположить, что это управляется на клеточном уровне. Но нет, клетки не могут сами по себе знать какого уровня проявления их внутренних программ реализовано на верхне-организменном уровне. Это регулируется посредством обратных связей между реализацией программ на верхнем уровне и соответствие их внутренним. Таким образом, это обоюдный процесс регулировки, осуществляемый сразу на двух противоположных полюсах.
На клеточном уровне это проявляется как в виде ограничения возможности клеток к полноценной энергетике, регенерационным и репаративным способностям, так и в виде перехода на новый высокий уровень выработки мортальных = смертельных провоспалительных веществ ведущих к отключению систем и тканей.
Можно утверждать, что старение есть выход за пределы оптимальных гомеостазов, интеграция которых обеспечивает коридор гормезиса, в пределах которого возможна нормальная реализация клеточных функции и их жизнедеятельности. При выходе за пределы гормезиса ткани работают не в рамках оптимума и это запускает в них программы провоспаления.
Старение является логическим этапом завершения не просто зрелости организма, а его перезревания, то есть нахождения уже за пределами оптимальных параметров как на общеорганизменном уровне, так и на клеточном. Выход за пределы оптимума осуществляется изнутри, когда уже нет очередных компенсаторных программ. Весь резерв программ развития исчерпан, а возвратных в исходное состояние нет, или потенциал их крайне слаб по отношению к противоположным программам на репарацию и регенерацию и реювенилизацию.
Может быть задача в том, чтобы найти способ сместить баланс соотношения в пользу первых сил по отношению ко вторым.
Интерлейкиновые провоспалительные вещества – механизм реализации запрограммированного старения.
Основной вред в механизмах старения приносят не какие-то токсины, а самые банальные клеточные гормоны воспаления – интерлейкины, которые в определенных количествах приносят пользу организму, а в крайних случаях при их избытке – огромный вред. Их избыток определяет как новый гормональный профиль, так и ограничение возможностей клеток из-за исчерпания возможностей стволовых. В новых условиях сенелита они уже проявляют не своё защитное и репаративное предназначение по запуску программ противодействия патологии, а выступают в роли гормонов смерти, которые выключают наш организм, отключают жизнедеятельность его важнейших органов: сердце, лёгкие, мозг… Это авральная программа на ускорение самоуничтожения, самоликвидации организма. Как результат именно последствий воспалительных интерлейкинов – гибель организма.
Усиление выработки интерлейкинов является частью неизбежной программы сенелита для мортального = смертельного исхода при всей неизлечимой хронике. Это механизм, задуманный природой по отключению организма, по частям отключения его важнейших систем жизнедеятельности. Это механизм завершения, отключения, несовместимости ситуации с дальнейшим пролонгированием жизни. По сути его можно сравнить с механизмом апоптоза клеток, но на уровне организма = самоликвидации организма. Дальше нет смысла поддерживать этот разваливаюшийся организм как систему систем. Это механизм, обеспечивающий мортальность для хронических безвыходных ситуаций. Природа не намерена поддерживать существование больных, изнемогающих организмов. Это специальный механизм отключения, элиминации. Всё тяжело больное должно быть элиминировано!
Итак, одни и те же механизмы проявления болезни имеется как у хронических нажитых заболеваний, так и у сцепленных со старением и являющихся его проявлением. Поэтому симптоматику старения трудно отличить от хроники.
И в том, и в другом случае организм может запустить механизмы самоликвидации, фенотипического апоптоза, то есть на уровне всего организма. У каждого вида живого может быть свой тип фенотипического апоптоза, различные механизмы его реализации. Оно завязано с онтогенетическими программами развития, то есть протягивания организма с одной фазы, этапа развития до другой, и, в конечном итоге, на фазе сенелита прорываются «шлюзы», когда программы провоспаления побеждают силы антивоспаления, идёт переполнение организма провоспалительными веществами, которые и обеспечивают лимитирование жизни. Механизмы противостояния провоспалениям исчерпаны. Интерлейкины обеспечивают фон, почву, на которой обязательно разовьются, вспыхнут буйным цветом мортальные болезни старости.
При старении такой шквал интерлейкинов уже не подавить теми способами, что достижимо еще при хронических заболеваниях. Этот интерлейкиновый шквал непреодолим. Это целый водопад, которому организму уже нет сил противодействовать, нечего противопоставить.
Основная причина ослабления защитных сил – это исчерпание возможностей стволовых клеток. Только ювенильные клетки способны выставить силы противодействия и репарации. У стареющих клеток такой возможности нет.
Это означает, что если нам удастся увеличить силы стволовых клеток, то мы сможем и дальше протягивать силы ревитализации и ювенилизации. Но эти силы закрыты переходом в новую комнату степени потентности. Проход в эту комнату полностью исключает возможность обратного выхода. Этому препятствует система генетических ограничителей = генетических блоков, не дающих передвигаться в обратную сторону по «реостату» генетических программ онтогенеза.
Топофизис – что это такое?
Какую роль он имеет в онтогенезе растений? В буквальном смысле это означает способность черенкованных клонов от дерева сохранять ту особенность роста у новых побегов от этих черенков, которой они обладали в кроне, например плагиотропный, то есть рост горизонтальный, как это делает ветвь кроне, или рост ортотропный – то есть рост вертикальный.
Циклофизис - что это такое?
Также клоны могут сохранять те особенности кроны, которые связаны с проявлением его генеративной готовности = зрелости побега, то есть, например закладки стробил у хвойных или цветочных почек у лиственных. В принципе эти понятия термины могут подменять друг друга, так как циклофизис не может проявиться в большинстве случаев без топофизиса, они обычно связаны.
Нами расширено понятие циклофизис и перенесено в физиологию животных организмов, где тоже имеется ряд аналогий. Циклофизис это онтогенетическая стадия проявления степени зрелости или особенностей взрослого морфогенеза побега. Она связана с изменением качественных свойств в меристемах почек, то есть аналога точек роста у животных.
Сохранение взрослых свойств у клоновых потомков не обязательно у всех видов растений, но у хвойных взрослых растений это довольно жесткая программа, когда многие последующие поколения клонов продолжают сохранять эти особенности их развития в кроне.
Циклофизисное старение.
Это когда клоны старых растений сохраняют признаки топофиза и циклофизиса, одного из этапов онтогенеза, в том числе и старости. Нами предложено пойти дальше и применить эти термины для животного мира. В этом случае мы подразумеваем возрастное сохранение признаков зрелости или старости как в стволовых клетках, так и тканей, которые они обеспечивают. От молодых клонов мы получаем качественно ювенильное потомство, а от старых клонов качественно сенелитное потомство. Казалось бы, в новых благоприятных для роста условиях сенелитное потомство должно реювенилизировать, но в практике этого не происходит. Внутренние программы необходимые для этого заперты.
Точно такая же ситуация происходит и в организме животных, когда меристемы или точки роста животных, подошедшие к этапу зрелости или перезрелости, воссоздают опять аналогичные себе, то есть стареющие линии, но никак не омолаживаются. Возврат в предшествующую «комнату» или этап адальтуса (начальная зрелость) или ювенила перекрыт в связи с их генетическим перепрограммированием, включением новых программ. Ключи от предыдущих программ не производятся, так эти программы заблокированы.
Топофизис старения - очень важное понятие, так позволяет создать более понятную и правильную концепцию механизмов старения, связанное с продвижением по ступеням онтогенеза. В своём труде я переношу ряд понятий из физиологии растений в область концепции онтогенеза животных. В этих вновь введенных понятиях четко просматривается суть старения тканей как у растений, так и у животных. Тем самым, мною сделано предложение пересмотра понимания биологического смыла механизма старения.
Концепции старения животных, которые увязывает её с некими теломерами, эффектом Хейфлика, эпигенетикой ведут к увязанию в тупиковом болоте, так не увязывает её с разворачиванием онтогенетических программ и продвижения по этой лестнице. Старение в царстве растений четко проявляется с продвижением по этой лестнице. Всё остальное вторично. Старение растения контролируется как на уровне всего организма, достижением необходимых заданных параметров, так и на уровне генов, которые определяют, как полноценно развернулся данный этап развития.
Реостат онтогенеза.
В этой работе мы рассмотрим, как происходит переход с одних генетических программ развития на другие по принципу реостата. На одном полюсе его находится комплекс ювенильных программ, а на другом – сенелитных. Без предшествующих реализаций практически невозможно включить последующие программы. Дело в том, что последующая программа, например зрелости реализуется не на уровне генов, а сверху, то есть на уровне достижения необходимых изменений, например количественных, на уровне организма, то есть регулируется целостным организмом. Пока эти внешние изменения не реализуются полностью, внутренние программы не могут переключиться. Причем переход на новую ступень онтогенеза подразумевает не нечто застывшую фазу, а её раскрытие, раскручивание, рост в определенных рамках, предусмотренных программой этого этапа. К тому же у разных видов программы на реализацию перехода в новую фазу развития различны. Например, у мыши все жизненные циклы осуществляются за 2 года, тогда как у голого землекопа он равен 33 года. Растянутые циклы онтогенеза определяются особой чувствительностью генных программ на переход к новому этапу.
Конечно, в мире живого многие промежуточные программы могут быть аннулированы или наоборот включены иные вставочные программы, которые и определяют особенности онтогенеза от короткоживущих видов, например у пчел – 56 дней, до мафусаиловых видов, которые могут жить несколько тысяч лет - секвойядендрон.
Сенелит или старение.
Тем не менее, термин старение лучше заменить на термин сенелит, так как он более четко подразумевает продвижение на следующую ступень лестницы онтогенеза, достижения очередной возрастной фазы развития, а не простое старение, например из-за плохого качества жизни.
Пример полиморфизма побегов в кроне секвойи как показатель степени их генеративной и онтогенетической разнокачественности.
Полиморфизм является хорошей подсказкой понимания работы генов в разные этапы развития и возможности их реювенилизации.
Для понимания вопроса приведу пример из моей научной диссертации в аспирантуре, когда разрабатывались способы вегетативного размножения секвойи. Проблема была в том, что зрелые побеги из кроны практически не дают способности к укоренению. Ихние гены не способны к включению на полную мощь программ регенерации, то есть отрастания корней, а если та незначительная часть черенков и образовывали корни, то саженцы были уродливы с образованием 1-2 слабых корешков и топофизисного характера роста, то есть сохранение тех особенностей роста, которые они имели в кроне: ограниченный, слабый темп роста как у ветви, преобладание плагиотропности = горизонтального роста, как растут ветви у хвойного дерева, а не вверх = ортотропно.

На фото: Различия укорененных черенков, взятых из различных частей кроны дерева секвойи.
Из фото видно, что побеги 1-2, взятые снизу кроны от плагиотропных ветвей практически не образуют корни, а внизу образуется каллусная шишка – нарост. Побеги под номером 3-4 взяты из средней части кроны и образуют промежуточное состояние между каллусообразованием и одиночными корнями, а под номером 5-6 побеги с приростом взятые из верхней части кроны от ортотропных ветвей и образуют более качественную корневую систему.
Образование каллусной шишки вместо корней в основании черенка связано со смещением гормонального профиля = смены гено-программного обеспечения = перепрограммирование, переход на иной комплекс программ данной части побега в нижней части кроны. Это переход на терминальные программы развития = крайние, дальше которых уже нет следующих, а есть только постепенное усыхание и гибель побега. Гормонального оживления меристем здесь нет. Апикальная точка роста побега заглохла. Это так называемое локальное старение побегов у растительного организма.
Следует признать, что аналогичные механизмы и у тотального старения всего растительного организма, когда доходит до того, что мощности, интенсивности гормонального фона недостаточно для нормальной работы главных апикальных почек.
У легко укореняющихся культур и у побегов из ювенильной кроны черенки не дают каллусную шишку, например у гортензии, тополя. Значит накопление ингибиторных гормонов у них минимально.
Гормональный фон апекса в свою очередь определяет состояние развитости или неразвитости камбиального слоя. Именно из камбия могут регенерировать корешки. При недоразвитом камбии даже если мы будем обрабатывать черенки ауксиновыми гормонами для стимулирования образования корней, то даже это не поможет из-за недоразвитого камбия и наличия гормонов стресса отменяющих ауксиновый стимул. В первую очередь здесь работают гормоны ограничители роста, а вернее абсцизовая кислота, которая в норме проявляет себя как гормон стресса, но в данном случает отвечает за зарастание раневой поверхности пробковой тканью, образующейся из феллогена. Ауксиновые гормоны, которые отвечают за регенерацию корней, а также гиббереллины – гормоны, которые отвечают за растяжку побега, здесь отсутствуют.
Подведем итоги: проведенные наблюдения четко показывают, как:
- Гормональный профиль, то есть на уровне целостного растения отвечает за переключение гено-программ. Сами они не могут включиться пока им не придет сигнал сверху.
- Двойственное = двустороннее = двуполюсное управление развитием: как сверху на уровне активности апикальных меристем и уровня гормонального профиля, так и на нижнем уровне гено-программ. Нижние гено-программы не переключаются на новые до тех пор, пока не проработается развитие на верхнем уровне, не исчерпается предел их возможностей. Эта проработка даст новый гормональный профиль и соответствующее ему апикально-камбиальное строение побегов и связанных с этим морфогенетическим типов побегов и почек.
Апикально-камбиальная активность растений аналог гормональной оси регулировки организма через эпифиз – гормональные железы мишени – точки роста во всех органах животного организма.
Если в растении апикально-камбиальная активность определяет онтогенетический статус растения, степень его зрелости, то в организме животного его определяет ось: эпифиз - гормональные железы мишени – ростовые точки тканей. Через ростовые точки определяются переходы в адальтус-матура-сенелит. Если сравнить хвойные деревья старение, у которых начинается в первую очередь на плагиотропных = второстепенных побегах, то у человека оно начинается на главной оси – эпифиза = аналога главному апексу и матрикальному = центральному побегу у хвойных. Это означает, что старение у человека организовано филогенезом так, что оно запускается через центральную ось, а она уже к 20 годам дает программу на сворачивание, ограничение жизненной силы организма. Возрастное развитие, а затем и сенелит определяет не клеточный уровень, а реализация неких верхних промежуточных программ становления, в частности адальтуса и матуры. Если стадии адальтуса и матуры как-то устранить, то и сенелит не подключится.
В этом случае организм застынет на стадии ювенила и будет ждать, когда придет клеточный черед ограничительных программ сенелита. В тоже время ювенил это своеобразный коридор, по которому идёт динамический процесс изменений в виде вегетирования, роста, для которых у организма всегда есть рамки, ограничители роста. Зрелость — это переход от количественных изменений к качественным. Любой ювенил имеет суживающийся коридор, в котором он не может находиться беспрерывно.
Если же эти закономерности перенести на организм животного, то здесь будет полная аналогия. С возрастом здесь также уменьшается как у растений активность камбия, так у животных точки роста и регенерации. Если в растении гормональный профиль определяет состояние апекса, то в организме животного – состояние эпифиза.
Но кроме гормональных перекосов здесь еще имеется слабый латеральный камбиальный слой = слой стволовых растительных клеток, который обеспечивает рост побега или растения. Камбием управляет апекс, какой апекс такой и камбий. Отсутствие или недостаточность изначально латеральных камбиальных слоёв тоже отвечает за слабый регенеративный ответ побега.
Итак, за переключение гено-программ на сенелит отвечают гормоны стресса – например абсцизовая кислота и этилен при практически полном отсутствии гормонов роста, деления и регенерации – например ауксины, цитокинины, а также мощность камбиальных слоёв. Отсюда прорисовываются существенные различия гормонального профиля как в стареющих растениях, так и в зависимости от расположения наиболее продвинутых по ступенькам онтогенеза побегов в кроне или у клоновых потомств, взятых от старых маточных растений.
Это означает, что не все побеги в кроне дерева имеют одинаковую степень онтогенетической зрелости. Проявляется возрастная разнокачественность их зрелости. На дереве имеются одновременно как побеги в стадии ювенила, так и адальтуса, матуры и сенелита. Но с увеличением возраста соотношение этих побегов увеличивается в сторону сенелита. Суммарный гормональный фон боковых ветвей дерева определяет степень активности центрального апекса. Это и определяет полный цикл развития древесной особи. Затухает рост главного матрикального побега и ведущего апекса, а вместе с ними затухает и камбиальная активность. Ослабление камбиальной активности ведет к ослаблению защитных свойств от грибков и паразитов.
Правда данная схема изменения гормонального профиля дана на примере хвойного растения секвойи. Но тем не менее, этот же принцип имеется и у растений, не имеющих центрального апекса и ствола, например кустарники, и у растений с коротким циклом жизни = 1,2,3 и т. д. года.
Возможные пути предотвращения выхода за пределы гено-программы ювенила.
Что бы этот «коридор» не сужался решение может осуществляться в природе несколькими путями:
- Путь самоклонирования. В природе известны самоклонирующиеся особи возраст которых исчисляется 11 тысяч лет.
- Самоустранение стареющих линий клеток животных с помощью их аутолиза или апоптоза.
- Усиление вегетативной фазы над генеративной.
Методики по управлению этими механизмами находятся пока в теоретической и практической разработке. В тоже время известно, что циклические голодания могут запустить механизмы аутофагии больных и слабых клеток, освобождающих место молодым и здоровым линиям клеток. Доказано что мыши при этом увеличивают продолжительность жизни на 40%. Но есть надежда, что можно пересечь и эту планку если специальными чередованиями длительных курсов голодания с кратковременными.
В тоже время следует понимать, что просто голодание является стрессом и несёт в себе кроме положительной стороны и негативную – истощение и ускорение включения механизмов сенелита. Палка о двух концах. Здесь следует уметь эквилибрировать между двумя крайностями. Этому и будут способствовать методы усиления вегетирующих стимулов, но на фоне ограничения калорийности = энергетической стороны питания.
Проводить реювенилизацию следует на фоне специальной витализирующей диеты с биогенными продуктами и специальными ростовыми стимуляторами, например королевское маточное молочко пчел, зародыши яиц, почки и побеги активно растущих ростков растений. При этом значительно ограничивается высококалорийные монопродукты на основе углеводов и жиров, но в тоже время усиливается приём высокобелковой пищи, лучше из морской рыбы, бобовых проростков и даже мозговые ткани животных.
Главный апекс растений и система эпифиз-гипофиз человека имеют одинаковую функцию – внешнего дирижёра как гормонального профиля, так и для процессов становления программ на уровне клеток и их геномов.
В организме человека аналогом апикальной точки роста или дирижёра развития можно признать систему эпифиз-гипофиз, которые и отвечают за весь гормональный профиль организма. С возрастом работа эпифиза затухает или прекращается полностью и происходит гормональный возрастной перекос, что и включает гено-программы матуры и сенелита, когда преобладают гормоны воспаления и стресса. Такие клеточные клоны в культуре ткани будут сохранять туже степень зрелости, продвинутости по реостату возрастных программ.
В свою очередь эпифиз закольцован с работой гипоталамуса и гипофиза. Все они обеспечивают нейрогормональное онтогенетическое развитие и соответственно их профиль.

На фото: 32-летний мужчина, который перестал стареть 20 лет назад и выглядит как школьник.
Такое отклонение в темпах созревания объясняют особенностями его гипофиза, а вернее недоразвитостью его. Возможно, здесь проявляется одна из форм гипофизарного нанизма, где имеется не только слабое выделение гормона роста, но и гонадотропинов, которые стимулируют половое созревание. Это в свою очередь ведет к недоразвитию вторичных признаков пубертата – одного из симптомов неотении. Тем не менее, сохранены первичные признаки пубертата, то есть оволоснение лобка и нормальные половые функции, в том числе либидо, оргазм и фертильность. Это говорит о том, что вторичный пубертат связан всего лишь с более высоким уровнем половых гормонов – тестостерона.
В свою очередь напомню, что высокие уровни андрогенов связаны со стадией адальтуса, а затем матуры и тем самым выводят организм за пределы коридора гормонального оптимума.

На фото: мальчик 8 лет с преждевременными признаками старения или прогерия.
Аналогичность программ и механизмов старения у растений и животных.
Камбиальные слои побегов и меристемные зачатки в почках можно считать аналогами точек роста и стволовых клеток в организме животных. С возрастом эти аналоги в организме животных уменьшаются по тем же самым причинам, что и в организме растений, где их включают программы на циклофизис = генеративное перезревание. Терминальная степень циклофизиса является началом сенелита растения. В организме животных терминальная степень адальтуса тоже является началом сенелита организма животного. Почему говорится терминальная степень, а не стадия адальтуса? Дело в том, что адальтус это не что-то застывшее, а постоянное продвижение по коридору. Рамки этого коридора, по которому ему нужно пройти, обусловлены гено-программами. Затем динамические процессы постепенно выходят за эти рамки, то есть за рамки оптимума и появляется необходимость запускать новые программы стабилизации и сдерживания, пролонгирования жизни в рамках онтогенеза. В обоих случаях как животного организма, так и растительного гено-программы предшествующих стадий развития глубоко заблокированы. Эта блокировка продолжается не только в клоновых потомствах, но и в клеточной культуре ткани. У растений эти блоки частично можно преодолеть в культурах ткани, меняя гормональный профиль и тем самым включить перепрограммирование и вызвать омоложение клонов. У животных эти блокировки на уровне гено-программ глубже и доходят до стадии зиготы и постэмбриональной диапаузы. Внешне это проявляется в ограниченном числе митозов, что описывается как эффект Хейфлика.
Можно утверждать, что у побегов ортотропного строения, более выражено апикальное доминирование, которое и определяет более высокий гормональный профиль, что в свою очередь определяет большие возможности побега к вегетационным программам, а заодно и к регенеративным возможностям. В свою очередь центральный верхушечный = матрикальный побег не проявлял способности к образованию корней, что свидетельствует об некотором сверхвысоком одностороннем гормональном фоне в этом побеге, тоже ограничивающем его возможности к регенерации.
Это означает, что гены ювенильности у таких побегов заблокированы даже в их клонах = потомках. Причём, чем более взрослое дерево секвойя, тем больше полиморфизм = разнообразие побегов по стволу. Особую плагиотропность проявляют побеги из нижней части кроны ствола, тогда как верхушечные ветви проявляли большую ортотропность, но при этом имели выраженную зрелость, то есть циклофизис в отношении образования мужских стробил = пыльников, тогда как ветви средней части кроны обладали потенцией к закладке женских стробил = шишек.
Обычно чем выше по кроне заготовлены черенки, или чем больше возраст маточника, тем более заблокированы гены ювенильности и выше степень проявления топофизиса у их клонов, тем выше у них степень появления побегов генеративного типа = способных к закладке шишек-стробилов, и ниже вегетативные = ростовые способности. Как видим, здесь постепенно подключаются новые программы созревания = адальтуса, но каждое очередное продвижение по программам зрелости отключает еще сильнее программы ювенильности.
Тотипотентностью у секвойи обладают только спящие = полностью ювенильные почки, которые не заходили в следующие «комнаты» = этапы развития.
Если побеги от спящих почек обрезать или содержать в темноте их основание, что называется этиолированием, то они способны перезакладывать аналогичные себе спящие почки и так до бесконечности. Как видим для сохранения или поддержания ювенильности ювенильным побегам нужны специальные условия, например полное отсутствие света. Тогда ювенильная почка запустит образование ювенильной. Свет как фактор развития переводит их на новые программы взрослого развития.
Ювенильные маточники.
![]() Гарбузов Г. А. показал в своей научной работе, что можно создать вокруг основания ствола огромную массу спящих почек и пучки из ювенильных побегов наподобие «ведьминых мётел», которые великолепно способны к регенерации корней = укоренению черенков. Главное, чтобы эти побеги из спящих почек не проходили стадию созревания, когда они вынуждены переходить в «комнату», где созданы условия для перехода на новые генетические программы. Здесь их ювенильная программа моментально отключается = блокируется. Причем блокирование настолько мощное, что даже многократное клонирование черенков от этих растений не позволяет им вернуться в первую «комнату» ювенильности.
Гарбузов Г. А. показал в своей научной работе, что можно создать вокруг основания ствола огромную массу спящих почек и пучки из ювенильных побегов наподобие «ведьминых мётел», которые великолепно способны к регенерации корней = укоренению черенков. Главное, чтобы эти побеги из спящих почек не проходили стадию созревания, когда они вынуждены переходить в «комнату», где созданы условия для перехода на новые генетические программы. Здесь их ювенильная программа моментально отключается = блокируется. Причем блокирование настолько мощное, что даже многократное клонирование черенков от этих растений не позволяет им вернуться в первую «комнату» ювенильности.

На фото: Внешний вид порослевого маточника для получения ортотропных ювенильных побегов для заготовки легко укореняемых черенков.
Маточник создан у основания ствола старого дерева из находящихся там изначально спящих ювенильных почек. Черенки из зрелых побегов, взятых в кроне этого же дерева, плохо укореняются.
Этиоляция – важный фактор ювенилизации у растений.
Сдует обратить внимание на особенности черенков, взятых от этих маточников: нижняя часть их ортотропно вытянута и этиолирована = полное отсутствие пигментов делает побег белым, а полное отсутствие ингибиторов позволяет таким черенкам легко регенерировать - укореняться. Этиоляция отдельный важный фактор для закладки, а вернее перезакладки новых именно ювенильного типа почек, то есть почек с ювенильной меристемой. Это своеобразная точка роста с сохранением ювенильной меристемы. Причем после обрезания-заготовки таких черенков на оставшихся необрезанных нижних частях может происходить закладка до бесконечности множества новых почек и отростков из них с ювенильными свойствами. Такие ювенильные маточники могут существовать теоретически вечно.

На фото: справа особенности черенков, заготавливаемых из ювенильных маточников с признаками этиоляции нижней части побега
Свет является фактором быстрого созревания растения и перехода его и его меристем = точек роста на новые этапы онтогенетического развития. Этиоляция = отсутствие света включает программы на снятие степени зрелости и переподключает на обратные программы отвечающие за облегчение способности к укоренению = регенерации, а значит к ювенилизации.

На фото: Наглядный пример обилия прорастания тотипотентных спящих почек у ювенильного порослевого маточника секвойи с образованием побегов с этиолированным основанием и фасциациями = срастанием у части побегов.
Анализируя особенности отрастания этих спящих почек, обладающих полной тотипотентностью, а также их бурную энергию роста, следует отметить, что из этих почек образуются густо новые побеги, необычным у которых является этиолированное основание, а также часть побегов образуют фасциации – срастания друг с другом. Фасциации связаны с бурным ростом и особым гормональным фоном, когда еще недостаточно факторов или гормонов для включения дифференцировочных программ, обеспечивающих механизмы разъединения сросшихся побегов. Для этого обычно нужно запустить программы апоптоза, а последний может запуститься только при определенных условиях гормональных соотношений или наличия нужных факторов, что сопряжено с включением генов дифференциации.
Новые побеги отрастают после срезания ранее побегов на черенки, а в оставшейся не срезанной нижней части образуется в пазухах хвоинок неограниченное ничем количество новых почек и сросшихся побегов с тотипотентными = ювенильными свойствами. Из фото видно, что из одной почки может вылазить целый пучок новых этиолированных побегов. Следует отметить огромную интенсивность и бурность отрастания новых побегов.
Для чего необходимо рассмотреть так подробно этот процесс? Здесь находится наводящий ответ на причины секрета ювенильности, суть которой кроется не просто в наличии определенного набора гормонов, их профиля, а именно в особо высоком уровне ряда гормонов, что обеспечивает не просто высочайшую энергию роста, но также переключает онтогенетические программы развития на ювенильные. Последние в свою очередь сопряжены с отсутствием веществ ингибиторов. Это означает, что вводится блокировка генов, ограничивающих рост.
В человеческом организме эту энергетику = потенциал тоже создают с одной стороны как высокий гормональный профиль, а с другой стороны задел потенциала клеточной способности к митозам, репликациям. Как известно, со временем репликативность резко падает, что связано с ограничительным феноменом Хейфлика.
Понимание механизмов рознокачественности клоновых саженцев, полученных из черенков от ювенильных маточников и из кроны позволяет объяснить ряд механизмов онтогенеза и сенелита, а также причины возможности или невозможности их обратимости.
На фото видна большая степень разнокачественности саженцев, взятых из ювенильных побегов, заготовленных в порослевом маточнике из спящих почек и их побегов и заготовленных в кроне того же дерева. Особенностью саженцев из ювенильных побегов является высокое качество корневой системы, что говорит о высокой способности к регенерации у них, а также анизофилия = отличие в строении лиственных побегов и вытянутости главного стволового побега, что говорит о наличии гиббереллиновых гормонов.
Саженцы из топофизисных черенков могут сохранять уродливый, угнетенный и боковой рост многие годы, а иногда и навсегда. В некоторой степени содействие по выходу из этого топофизисного состояния может оказать приём по чрезмерному удобрению азотными удобрениями, что приведет к усилению вегетативной фазы и «взлому» гено-программ.
Обратите внимание на этот приём «взлома» путём активации вегетативной фазы, что приводит к реювенилизации = переключению программ в обратную сторону. Следовательно, в принципе это возможно в мире растений. В дальнейшем мы покажем, что это возможно и в мире насекомых, когда погибшую неотеническую матку заменяют обычные особи, например у термитов, а также в мире животных. Этот же прием активации можно и нужно осуществить и в организме человека.
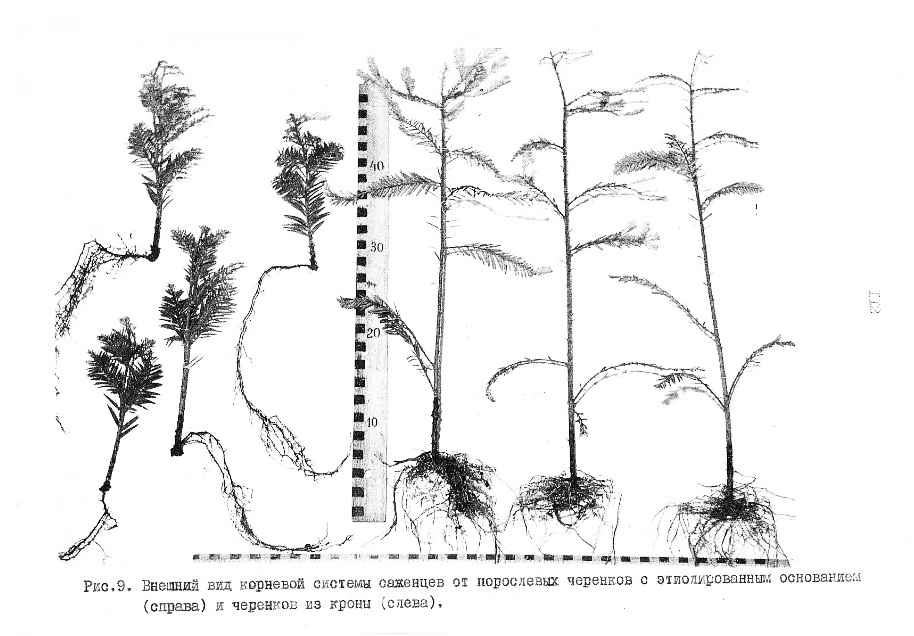
На фото: справа внешний вид укорененных черенков-саженцев от ювенильных ортотропных побегов, заготовленных в ювенильном маточнике, которые сохраняют все признаки ювенильного строения сеянцев и высокой степени к регенерации. Слева укорененные черенки от побегов, взятых в кроне.
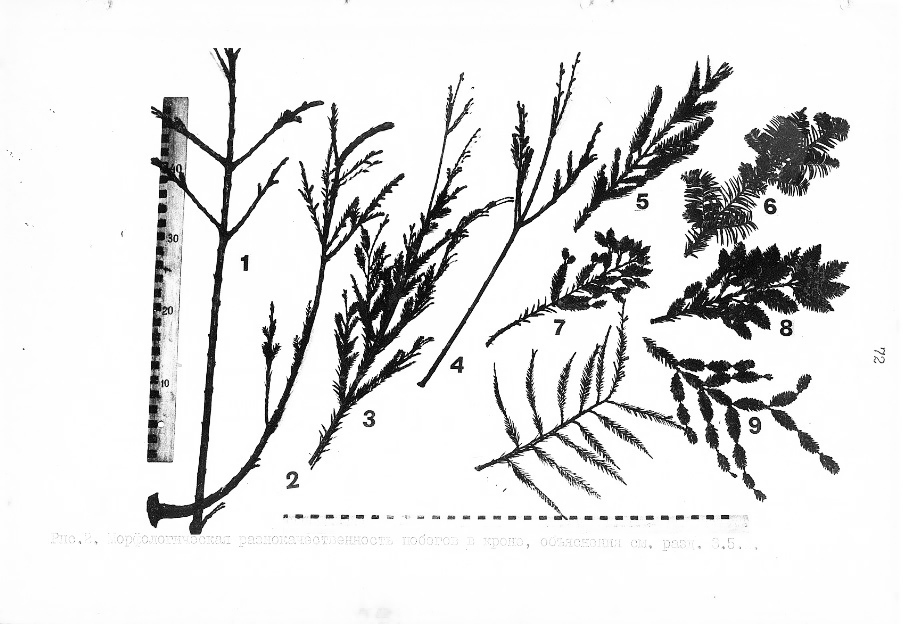
На фото: видна широкая морфологическая разнокачественность побегов в зависимости от их расположения в кроне.
На фото представлено морфологическое разнообразие и типы побегов в зависимости от уровня их расположения в кроне и угла наклона ветви по отношению к стволу.

Выяснено, что на морфологическое строение побега у секвойи влияет геотропизм ветви (угол наклона) и порядок ветвления и мутовок.
Все эти факторы определяют и особенности гормонального профиля – фона в данном участке побега. То есть в различных частях кроны дерева гормональный профиль = соотношение спектра различных гормонов различно. Этот фон гормонов и определяет морфогенез побега.
Как видим нижние агеотропные побеги (направленные вниз) и высшего порядка ветвления обладают минимальной способностью к регенерации = укоренению черенков, а значит обладают высшей степенью циклофизисного старения. Но в этом термине не подразумевается старение в смысле изнашивание, одряхление, а старение, связанное с гормональными перестройками и блокированием всех более ювенильных программ, ведущее к максимальному угнетению роста побегов и способности к регенерации. Обычно такие побеги отмирают, усыхают в кроне.
Геронтологическая концепция сенолитика.
В геронтологии появилась новое учение сенолитика, выдвигающую концепцию старения, связанную с накоплением стареющих клеток, когда в итоге ихнее действие превалирует над процессами омоложения и репарации. Эти стареющие линии выделяют вещества, которые отрицательно влияют на окружающие клетки. Поэтому в науке разрабатывают лечебные вещества – сенолитики, которые способны влиять на стареющие линии и вести к их анулированию.
Концепция старения как проявления циклофизиса.
Эта концепция очень много имеет сходств с нашей концепцией циклофизисного типа старения, когда в кроне накапливается все больше и больше ветвей плагиотропного и циклофизисного типа. Избыток их угнетает ортотропные побеги и развитие дерева в целом. Меристемные точки роста или почки от них проявляют все признаки старения, которое невозможно разблокировать в потомстве клонов.
Это дает основание считать, что суть этих концепций одинакова. Но только в ракурсе того, как мы видим эту проблему можно утверждать, что сенелит не неизбежен и его можно разблокировать, тогда как концепция сенолитиков понимает все однозначно, не видит генетической глубины регулировки развития и утверждает, что сенелит неизбежен и бороться с ним можно только путем уничтожения линий клеток обладающих сенелитическими свойствами.
Почки – меристемные точки роста, которые определяют как разнокачественность будущих из них побегов, так и степень циклофизисного старения.
На фото, что представляет различные типы побегов в кроне, видна их высокая степень разнокачественности. Все эти побеги дают и различного типа почки, которые различаются как по физиологическим свойствам, так и по внешним морфологическим различиям.
Самый крупный побег на фото №1 – это центральный = осевой = матрикальный даёт самую крупную почку-меристему. По сути, эта почка является дирижером, относительно которого идет корректировка роста и развития всех остальных побегов. Здесь находится главная апикальная точка = главный апекс. От неё зависит качество многих других типов почек.
На фото под №9 виден плагиотрапный побег имеющий самый маленький прирост и самые маленькие почки. Если присматриваться к этому побегу. То можно отметить, что на нём образуются новые побеги, которые меньше предыдущих. Это говорит о затухании такого побега и о том, что в скором времени он полностью прекратит рост, а затем и отмирать.
Здесь можно говорить о высшей степени проявления циклофизиса и связанного с ним циклофизисного старения данной как меристемной точки роста, так клонов из него. Из этого вывода следует другой вывод, о том, что уровни старения точек роста в кроне дерева различен и максимален циклофизисный тип старения у самых нижних побегов в кроне.
Итак, мы переходим по «отсекам коридора» = гормезиса, где на первой ступени возможна полная безграничность, бессмертность = иммортальность, а в последней комнате запускаются механизмы мортальности = самоликвидации за счет самосжигания в пламени провоспаления.
Напомню, что ювенильные программы возможны только на начальных этапах онтогенеза особи или растения. Каждая программа развития обеспечивается за счет вначале тотипотентных стволовых клеток, из которых затем происходит вход в комнату, где реализуется новый уровень регулировки с помощью плюрипотентных, затем идёт переход в комнату, где правят мультипотентные клетки, а затем - в унипотентные клетки и т. д. Клетки шишек находятся на ступени монопотентности и способны к дальнейшим делениям, а направлены только на путь самовыключения, отмирания. Уточню, что простое отмирание ткани не является апоптозом, когда гибнущие клетки исчезают, саморастворяются. Тотипотентные программы заблокированы на самом раннем этапе развития растения, только они способны дать зачаток побегам с ювенильными свойствами. Именно эти побеги и черенки из них способны к максимальной регенерации и восстановлению корней, причем равноценных по качеству сеянцам этого растения. Таким образом, растительная особь проходит по лестнице самых различных программ развития = онтогенеза.
Итак, особенностью секвойи является уникальная способность легко закладывать «спящие» почки в основании ствола, которые обладают тотипотентными свойствами = ювенильными, которые не зависимо от общего возраста растения, даже 100 и 1000 лет, не теряют и не блокируют свои гены ювенильности. Причем спящие почки способны давать начало множеству новых спящих почек вплоть до бесконечности.
Из всего этого сказанного можно отметить, что существуют некие программы ограничители, которые строго перекрывают обратный проход в ювенильность. Очевидно, работают они за счет репрессии генов.
Нечто аналогичное происходит и в клетках человеческого организма. Зрелые ткани животных сохраняют свою зрелость = онтогенетическую продвинутость аналогично побегам-ветвей дерева, которые сохраняют свои морфологические особенности строения в зависимости от расположения в кроне даже в своих клонах. Это так называемое проявление топофизиса, то есть утраты способности к переключению на прежние или другие программы морфогенеза. Не пускают гены-ограничители. Омолодить их практически невозможно.
Но выход есть – это подать в организм тотипотентные белки-ключи, переключатели. Может создаться ложное впечатление, что тотипотентные белки запустят в тканях организма образование множества неких эмбриональных зачатков, нечто подобное на тератомы, то есть множество зачатков новых органов в новых местах. Но нет, их пробуждающую первичную инициацию, индукцию быстро откорректируют программы окружающих клеток, которые повернут инициацию в свою сторону по появлению новых эмбриональных клеток, но именно той ткани, в которой они находятся, из которых состоит орган или ткань. То есть быстро переходит переключение на плюрипотентные программы и т. д.
Главное обойти гены ограничители, сделать запуск-индукцию предшествующих программ.
То, что такое переключение программ возможно подтверждают данные по так называемым «ведьминым мётлам», когда на зрелых побегах появляется множество новых ювенильных побегов, растущих в несоответствии с программами морфогенеза. Причиной этому могут быть грибки, которые могут выделять стимулятор роста, например ауксины, возможно в неадекватных количествах, ломающих баланс гормонов в сторону ювенилизации, когда затронут механизм повышенной закладки меристем, но при этом нарушен морфогенез, который, по сути, и является одной из ограничительных программ возврата к прежним программам. Очевидно, стимулирующим фактором здесь является не сам гормон, а его количественное содержание. Для ювенильных побегов характерен особо высокий уровень этого гормона. Здесь похоже концентрация этого гормона обуславливает тотипотентность, а не некие специфичные тотипотентные белки – генные ключи. Можно ли эту идею, концепцию значимости концентрации индукторов перенести на организмы животных? Концепция имеет определенную логику. Ясно одно, что у стареющих организмов резко падает уровень ведущих гормонов мелатонина и серотонина.
Итак, эффект связан с отключением ограничителей и запуском предыдущих программ. Возможно, имеют место и мутации или другие факторы, которые отключают программы ограничители и запускают программы ювенилизации. Как видим, здесь осуществляется возврат в «предшествующие комнаты» и нарушается закономерная последовательность онтогенеза. Следует отметить, что ювенильные побеги и спящие почки здесь закладываются именно из зрелых побегов, а не из спящих и сохраняющих ювенильность почек, как у секвойи.

На фото: «ведьмина метла» с четко выраженным скоплением ювенильных побегов.
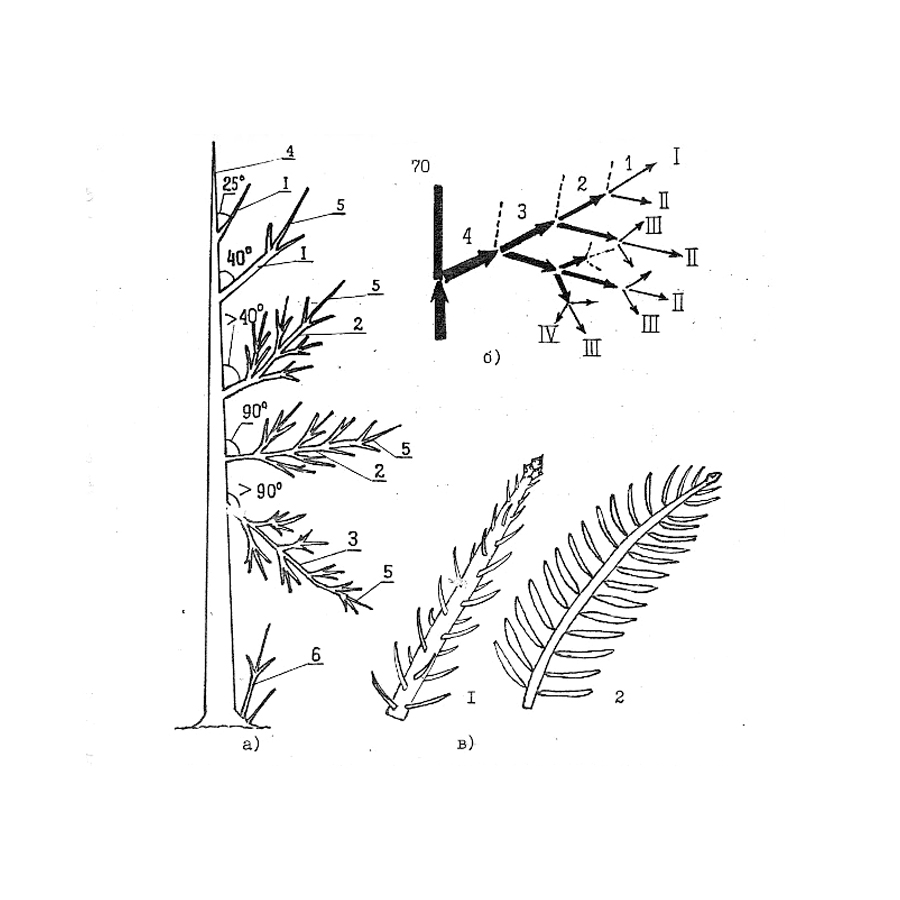
На рисунке: Расположение разнокачественных ветвей и побегов в кроне.
а). Морфологическая разнокачественность в зависимости от геотропизма ветвей:
1 – ортотропные, растущие вверх под углом 0-250 и до 400; 2 – плагиотропные, растущие под углом 40-900;
3 – агеотропные, растущие вниз;
4 – моноподиальный побег, лидирующий, образующий ось ствола;
5 – терминальные, осевые побеги скелетных ветвей;
6 – порослевые.
б). Разнокачественность побегов в скелетной ветви в зависимости от порядков ветвления (I, II, III, IV) и мутовок (1, 2, 3, 4).
в). Внешний вид разнокачественных побегов:
1 – с радиальным расположением хвои;
2 – в одной плоскости или дорсивентрально.
Любой морфологический тип побегов определяется соотношением не только тех или иных гормонов, но и их концентрацией. Из всего этого видно, что у растений как морфология, так и онтогенетическая зрелость определяется сверху – гормональным профилем, но в рамках генетических программ.
Здесь наблюдается замкнутый круг прямых и обратных связей: верхний гормональный уровень регулировок определяет контроль за генетическими программами, их переключение, а последние определяют рамки возможностей, коридор, по которым может пойти развитие во внешнем проявлении. Всё осуществляется, едет по рельсам, которые обеспечивают гено-программы, но реализуют их гормоны.
Здесь мы приблизились к самому главному вопросу этой книги: откуда исходит начало старения. Ответить, что сверху или снизу – будет неправильно, так все они закольцованы и выступают как одно единое. Если бы не было ограничивающих моментов, то рост мог бы происходить неограниченно, как это наблюдается у самоклонирующихся растений, например креозотового куста (Larrea tridentata), возраст колонии которого оценивается в 11700 лет. Но в лице одной особи этого достичь невозможно, так как рост такого растения-дерева сам себя ограничивает как в ширину, так и в высоту. Растение переключает механизмы перехода из вегетативной фазы в генеративную. Там, где начинают включаться механизмы ограничения роста, там запускаются механизмы адальтуса, а затем матуры и сенелита.
Сенелит может быть как запрограммированным организованным процессом на самоисключение, самоотключение, так и не запрограммированным и происходить произвольно, естественным путём, когда особь выходит из оптимального коридора существования и внешние факторы становятся сильнее для растения и добивают его постепенно. Внутренних программ противостояния у растения этим факторам больше нет, они исчерпаны. Жизненные силы становятся слабее внешних факторов. Поэтому старение не обязательно проявляется в виде программы, часто оно является следствием отсутствия программы.
Так медленно стоя, отмирают деревья гиганты. Конечно, старение завязано с хронологией жизни растения, его этапами. Скорее всего правильнее ответить – это динамический процесс, который раскручивается как снизу, так и сверху, когда идёт движение по ступеням реализации комплекса программ развития, но на определенном этапе хронологии входит в зону неоптимального существования, когда оно может быть обусловлено как изнутри генетически, так и внешними факторами.
Различие апикальной активности по уровню расположенности в кроне дерева.
Даже в пределах одного древесного растения на различных уровнях кроны апексы различны по степени тотипотентности. Чем ниже побег на кроне, тем больше они угнетены, слабее апексы. Самую высокую энергию тотипотентности дают меристемные клетки апексов из проростков семян, например орешков.
Правомочен вопрос: являются ли гено-ключи растений подходящими для животного мира?
Определённые, правда косвенные, доказательства этому есть. К сожалению, экспериментов, где использовались бы сугубо апексы в больших количествах, для лечения или оздоровления, в литературе не найдены. Но тем не менее, много материалов по применению для этих целей просто ростков растений, когда удавалось бороться с диабетом, ожирением, аллергией…
Мы питаемся преимущественно плодами, листьями, побегами, семенами находящихся в фазе покоя, в которых нет апикальных меристем, но зато много веществ ингибиторов, то есть с полярно противоположным действием. Ждать от них желаемого эффекта по ревитализации не приходится. Эти же ингибиторы не безразличны для нас. Они ослабляют нашу Жизненную Силу, ослабляют наши программы на ревитализацию. Один полюс ростка нас оживляет, а другой тормозит. Мы же в своей повседневной жизни используем только те части растений, которые имеют противоположный полюс действия. Пока мы молоды, нам хватает своих жизненных сил, то есть внутреннего потенциала, заложенного в нас от рождения, чтобы противостоять натиску противоположных сил, но с возрастом наш потенциал жизненных сил слабеет и его надо поддерживать.
Конечно, настоящего тотипотентного или подобного эффекта от этих апексов ожидать не следует, так как у растений иные гено-ключи чем у животных, но мощного оздоровительного эффекта можно ждать. Дело в том, что эти же вещества по своей природе более родственны для нашей микрофлоры кишечника. Здесь не должно быть по сути лимфоцитоза, гниения и брожения если пища переваривается преимущественно с помощью аутолиза. Только здоровый микробиом = микрофлора кишечника даёт гарантии нашему максимально высокому иммунитету = здоровью. Если в кишечнике нарушен микробиом и возбуждена лимфоцитарная защита, то это пролог к провоспалительному хроническому статусу организма – фундамента для многих неспецифических состояний, то есть ослабления и предрасположенности ко многим вялотекущим хроническим болезням. А это в свою очередь является истощением ресурсов наших защитных сил, изматывания иммунки. Это же является компонентой хронической усталости и преждевременного старения.
Таким образом, лучший союзник для нашей естественной микрофлоры – это наличие большого количества ростков, а лучше в виде апексов. Тогда мы не будем активно зря кормить 2-3 кг нашей микрофлоры, меньше будем затрачивать калорий и продуктов, а самое главное будет исключено ожирение и хронический провоспалительный статус.
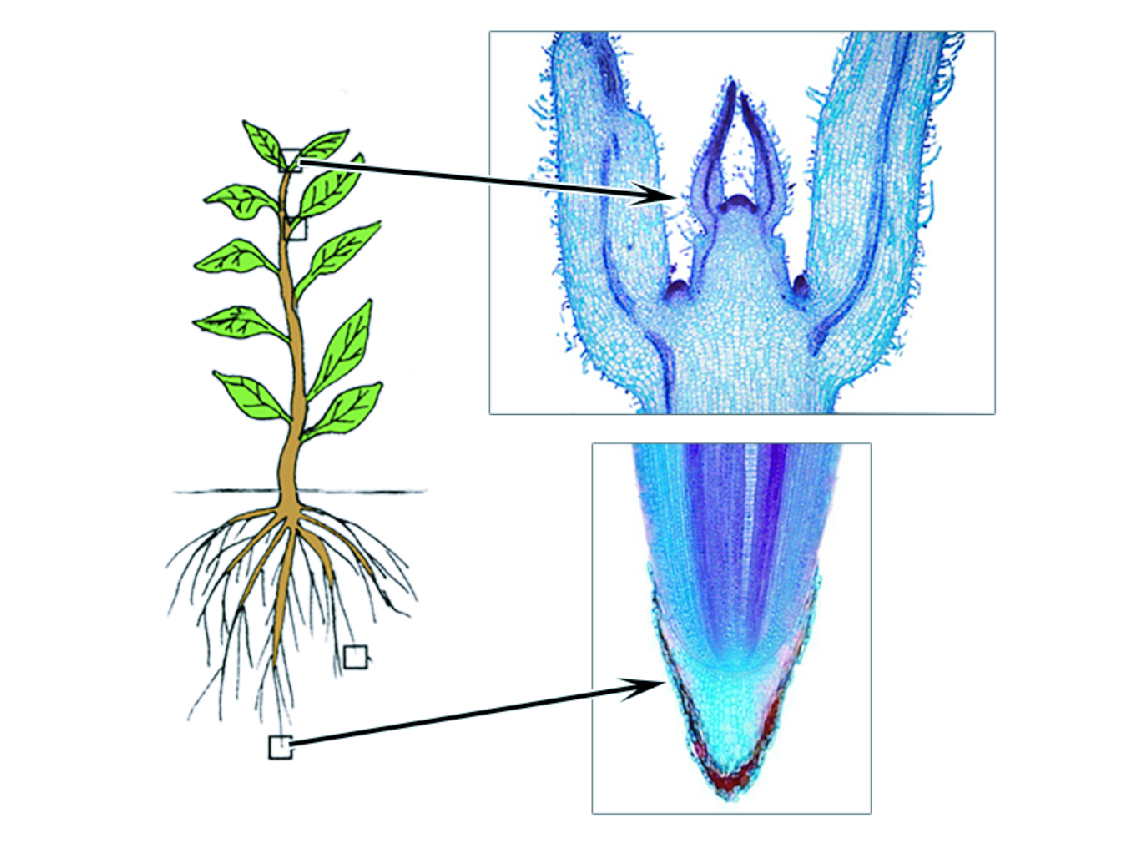
На рисунке: место расположения апексов.
Что означает регулировка программ на уровне целостного организма?
Нам более привычно, что программы регулируются на уровне клеток. В действительности регулировка идёт с двух сторон. Клетки просто анализирует в какой степени идёт продвижение по реализации в заданной программе. Например, программа роста = вегетации на заданном этапе развития организма имеет определенные рамки коридора, в которой она может реализоваться.
Коридор возможностей роста может быть очень большой и быть растянутым на многие годы, а может быть очень маленьким, например однолетний цикл развития. Все эти коридоры и определяют гены. Это так называемый клеточный уровень регулировок. У растений они обычно регулируются гормональным профилем = соотношением тем или иных гормонов. Период изменения этого профиля может быть очень растянутым и реализоваться вплоть до десятилетий. Это и есть границы одного из этапов онтогенеза.
Но организм не находится при этом в состоянии статики, то есть, например постоянно растёт, изменяет пропорции. На определенных фазах роста этот этап приходит к некоторым границам, ограничивающим непрерывный рост. Эти рамки границ тоже записаны на генетическом уровне. Как итог, это является сигнальным фактором, стимулирующим подключение новых программ развития, например ограничения вегетативного роста. Это и есть механизм перехода = переключения с одного этапа онтогенеза или развития организма на новый этап.
Шесть этапов постэмбрионального развития.
Автор считает правильнее выделять эти этапы под указанными терминами:
- Фетальный – У растений молодые ростки, а у людей грудничковый период.
- Ювенильный = натальный – детство и юность без признаков пубертата = половой зрелости.
- Имаго = постнатальный – Полностью развернутые программы морфогенетического становления организма, например у людей это соответствует подростковому периоду, но не с полным проявлением половых признаков, например отсутствие бороды = неполный пубертат.
- Адальтус – Имеет начальную полезную для организма функцию зрелости = полный пубертат, это, по сути, более повзрослевший этап развития, у людей это около 25-50 лет.
- Матура – Стадия перезрелости организма, например у растений это переизбыток закладки генеративных почек, резкое или полное торможение вегетативной мощи. Таким образом это генетическая программа на чрезмерность, идущая особи во вред; а у людей это совпадает с постклимактерическим периодом, связанным с резким изменением гормонального профиля, что ведет к ограничению генеративных возможностей в связи с выходом за пределы оптимума = коридора гормезиса для этой функции. Это ведет к резкому ослаблению Витаукта = Жизненной силы. Такая программа, как и прогерия = досрочного старения, которые ведут через разные механизмы, но к общему знаменателю - самоликвидации.
- Сенелит – Стадия появление как первых признаков старения, обозначаемых как патологическая врожденная синдроматика старения, сцепленных с генетическими программами, так и сопровождающего его Симптомокомплекса Болезней Старости (СБС), то есть приобретенных на фоне гормонального дисбаланса и клеточно-тканевых дисфункций. Сенелит с одной стороны является врожденным патологическим процессом, а с другой стороны на него накладывается СБС и у каждого индивида он специфичен и зависит от тех патогенетических факторов, с которыми организм встретится. Сенелит это одновременно врожденная патология = болезнь, а с другой эта же патология предрасполагает к комплексу других проявляющих болезнь факторов, которые не обязательны на жизненном пути, а в более молодом возрасте эти проявляющие болезнь факторы чаще всего не срабатывают благодаря более высокому противодействию Витаукт = Жизненной силе.
Именно переход на новую программу, то есть достижения целостным организмом этого состояния, и обеспечивает на общеорганизменном уровне возможность перехода на новые программы. Без этого клетки сами не могут запустить эти программы.
Принципы включения-выключения регулировочных программ онтогенеза.
На разных стадиях развития зародыша преобладает производство различных генных ключей. На первом этапе, самом начальном, преобладает производство генных ключей в виде тотипотентных белков, которые открывают первый замок развития, когда открывается дверь в комнату, которая является цехом по производству первичных стволовых клеток. В свою очередь ключами-индукторами для этого являются мультипотентные белки. Они-то и запускают процесс становления развёртывания бутона из трёх основных лепестков: эндо-, мезо- и эктодермального происхождения, которые и являются основой для становления всех последующих тканей. Каждый лепесток является особой комнатой-цехом для производства последующих генных ключей-индукторов – плюрипотентных белков, которые затем открывают замки для унипотентных белков. Такая вот лестница последовательного развёртывания программ развития зародыша и закладки всё новых и новых уровней и стадий становления последующих линий стволовых клеток.
Различие действия тотипотентных веществ в эмбрионах и во взрослом организме. При этом следует понимать, что те же вещества-индукторы = тотипотентные и другие будут по-разному действовать на клеточные массы в эмбриональной фазе развития и во взрослом состоянии проявленной дифференциации. Здесь уже их возможности будут заблокированы последующими программами. Нужно снять дифференциацию, то есть разблокировать эти клетки, чтобы запустить включение тотипотентных программ.
Все эти комнаты-программы раскрываются в яйце в течении 3-6 дней. Причём на каждый последующий день появляются всё новые и новые наборы ключей. Причем каждая открытая новая комната может блокировать работу предшествующей. Так производство первых тотипотентных ключей тормозится уже на самой ранней стадии диапаузы эмбриогенеза, на стадии зиготы. Это означает, что их производство закрывается уже в первые дни пробуждения зародыша.
Кроме того, чем дальше по коридору в следующую «комнату» идёт продвижение, тем больше программ включается на перекрытие всех предыдущих, тем меньше возможностей на универсальность возможностей. Это так называемое коммитирование – постепенное ограничение возможных направлений развития клетки. Коммитирование совершается ступенчато. Сначала соответствующие преобразования генома касаются крупных его участков. Затем всё более детализируются, поэтому вначале детерминируются наиболее общие свойства клеток, а затем и более частные.
Именно коммитирование обеспечивает возможность удерживать нужный дифференциал клетки, то есть включать высшие программы. Таким образом, одни программы жестко отключаются для того, чтобы дать возможность включиться новому набору программ. Дифференциальные программы в тотипотентном состоянии клетки = зиготы отключены = коммитированы, но открыты для тотипотентности. В свою очередь у зрелых клеток отключены, закрыты программы тотипотентности, но детерминированы = открыты программам детерминации. Это так называемая детерминация, то есть появление у клетки генетической запрограммированности только на один путь развития.
Коммитирование (открытие) детерминирование (закрытие) - два противоположных процесса, генетические программы, одна из которых исключает другую.
Этот коридор в цепочке между коммитированием и детерминированием можно сравнить с дроссельной системой управления программами развития клетки, где все программы записаны в определенной последовательности, очередности, но их надо запустить. Таким запуском, стартером является ползунок, наличие которого превращает дроссель в реостат.
Напомню, что реостат — это электрическая катушка с намотанной витками проволокой, по которой ездит ползунок. В клетке таким «ползунком» являются особые белковые факторы, например тотипотентные и другие. Причём «ползунок» здесь включает не один ген, а целый блок взаимодействующих программ.

На фото: реостат, принцип действия которого подобен переключению на новые гено-программы клетки при передвижении ползунка
Используя в пищу перезрелые семена, листья, мы способствуем всему комплексу процессов детерминации и дифференциации, усиливаем это крыло, ведём процесс прочитывания программ в сторону сенелитирования. Используя апексы и зародыши, мы инициируем коммитирование, то есть крыло программ в сторону реювенилизации.
В то же время наличие тотипотентных ключей является самым ценным началом инициации живого. В культурах ткани бесконечно могут делиться только клетки зиготы = тотипотентные. Тогда как на стадии бластулы они уже превращаются в плюрипотентные и способны делиться ограниченное количество = 60 раз. Это известный специалистам эффект Хейфлика. Жизненная сила таких клеток, как видим, намного слабее. Таким образом, яйца 2 – 3 – 4 – 5-дневного развития будут качественно отличаться по производству различных ключей.
Чем от более от ранней комнаты мы будем использовать ключи, тем глубже будет эффект реювенилизации от их применения. Естественно, лучший эффект дадут тотипотентные белки. Они самые ценные и они должны превалировать в предлагаемой нами смеси.
КЛЕТОЧНЫЙ УРОВЕНЬ СТАРЕНИЯ.
Проанализируем не является ли клеточным первичным старением примеры возрастного функционального снижения чувствительности рецепторов на мембранах клеток, которое приводит к снижению ими функциональной работоспособности, а также всегда сопряжено с митохондриальной недостаточностью. Последнюю увязывают с теломерной концепцией старения клеток, когда кончики хромосомных колпачков постепенно укорачиваются. Явление можно сказать присущее для всех типов тканей.
Таковые имеются, например на мембранах гормоночувствительных клеток простаты, что ведёт к первичному снижению их чувствительности и вторичной необходимости чрезмерного повышения уровня гормонов в крови. Также можно говорить и о возрастной толерантности клеток мышц к усвоению глюкозы. Можно сравнить толерантность диабетическую с толерантностью атеросклеротической или эпителиальной дисфункцией. И такие аналогии можно продолжать дальше. Везде среди каждого вида функционирующих тканей наблюдаются изменения на клеточном уровне. Можно ли их сравнить с первичным истинным старением клеток не определяемы верхними уровнями развития организма? Это в свою очередь может быть результатом как возрастного ухудшения питания ткани кислородом из-за вырождения, то есть возрастного уменьшения микрокапиллярной сети, так и следствием возрастного естественного старения клеток, ослабления их энергетики. Определенную роль здесь играет и изменение возрастного гормонального профиля. Всё это создаёт путаницу в понимании истоков, первоначал происхождения старения ткани: с верхних этажей или нижних. Правильность ответа на этот важный теоретический вопрос позволит искать правильные подходы в разработке принципов и методик противодействия этим пагубным процессам. Рецепторами обладают практически большинство типов клеток организма. То есть с возрастом снижается или теряется сенсорная чувствительность высоко функциональных клеток. Рецепторы взаимодействуют с сигнальными системами регулировок. Утрата рецепторов сопряжена с энергетическими проблемами = митохондриальной недостаточностью, а значит ослаблением клеток. Является ли это первичным клеточным уровнем старения? Отчасти да. Этому могут способствовать как как внешние для клеток факторы, которые им обеспечивает организм, а также факторы, исходящие изнутри клеток, например в связи с эффектом Хейфлика.
Где начало сугубо клеточного теломеразного старения, преждевременного старения и старения связанного с онтогенетическим стадийным становлением организма? Кроме механизмов старения как проявления онтогенетических этапов развития существует еще и более первичный уровень – старения на уровне клеток. Кто из этих уровней старения более важен для организма отдельный вопрос. А здесь рассмотрим в чем суть клеточного уровня.
Об эффекте Хейфлика следует поговорить отдельно.
Особенности стареющих клеток.
Доказано, что молодость клеток всегда проявляется в способности синтезировать группу химических веществ, называемых нуклеотидами, которые необходимы для формирования ДНК. Стареющие клетки их не производят. Старение сцеплено с потерей клетками потенции к размножению и это необратимое состояние клеточного цикла. В этом они проявляют полную противоположность стволовым клеткам. Мало того, так еще стареющие клетки с накоплением своей массы подавляют работу стволовых. Баланс взаимодействия между ними идёт в пользу стареющих линий.
Считается, что стареющие клетки никогда не смогут делиться. Но все-таки можно предполагать, что эта потенция закрыта в связи с переходом на особые терминальные этапы развития клетки, и это запрограммировано. Но эти программы можно снимать. Это, например можно увидеть на примере желтеющих листьев, которые обрабатывали гормоном цитокинином и их молодость и зелёный цвет восстанавливались.
Именно с такой ослабленной потенцией связывают множество болезней старости, например артрит, остеопороз, сердечно-сосудистые заболевания. Эксперименты показывают, что если у молодых клеток остановить производство нуклеотидов, то клетки стареют. Таким образом, производство нуклеотидов необходимо для сохранения молодости клеток и, если можно предотвратить потерю, клетки могли бы стареть медленнее. Именно через возобновление работы механизмов по воспроизводству нуклеотидов оказывает свое действие цитокинины на пожелтевших листьях.
В то же время ученые ищут возможную связь старения с механизмами, перекрывающими возможность проявления раковых перерождений. Тем не менее, со своей стороны, считаю, что это тупиковый подход поиска значения механизмов старения. Известно, что во многих случаях, например растениях мафусаилах, клоны которых могут размножаться веками, показывает, что старение здесь ни причем. При необходимости механизмы старения природа может легко обойти. Суть в том, что механизмы старения нужны как с филогенетических позиций эволюции, так и с позиции необходимости элиминировать неактивные части и ткани организма.
Не старение на клеточном уровне сдерживает рак, а наоборот старение на общеорганизменном уровне повышает его возможность. Ученые считают, что если нам удастся найти способ предотвращать превалирования стареющих линий клеток, то тем самым мы сможем предотвратить многие возрастные заболевания, а также увеличить продолжительность жизни особи. Поэтому выдуться работы по поиску методик и препаратов по удалению избыточных стареющих линий в организме.
Что представляют собой тоти-, плюри- и т. д. потентные вещества?
До сих пор ответа на этот вопрос отсутствует. Их можно предполагать, как некие белки, которые открывают первую универсальную программу зародыша. Только после открытия этой программы становится возможным открыть дорогу всем 350 программам по становлению клеточных зачатков тканей любого из них. То есть программы открываются пошагово.
Факторы, которые определяют уникальность стволовых клеток находятся не в ядре, а в цитоплазме. Это избыток мРНК всех 3 тысяч генов, которые отвечают за раннее развитие зародыша. Это означает, что все 3 тысячи ключей в виде мРНК находится заранее в цитоплазме. Следовательно, тотипотентными свойствами обладают не белки, а нуклеиновые кислоты.
Также из этих данных следует, что тотипотентными и плюрипотентными веществами индукторами является не одно вещество, а их целый комплекс. Тогда проходя через каждую «комнату» становления, они отсекают какие-то ключи, выбрасывают их из связки, а «комнату» блокируют. Но эти ключи отвечают за морфогенез, а нас интересуют программы онтогенеза.
То, что эти индукторы стволовых клеток находятся не в ядре клеток, а в их цитоплазме подтверждает эксперимент, когда ядро из соматической клетки пересаживают в овоцит оно теряет свою степень дифференциации = дедифференцируется = перепрограммируется и из него может вырасти полноценная взрослая особь.
Эмбриональные стволовые клетки = ЭСК. Также доказана возможность восстановления тотипотентности у соматических клеток путем простого слияния соматических клеток и ЭСК.
Что означает дифференцируется? Мы видели, что это возможно, например при пересадке ядра в цитоплазму стволовой клетки. Значит там находятся ключи. Куда исчезают эти ключи-индукторы после запуска зиготы и превращения в бластомеры?
Феномен хоуминга очень важен для понимания адресности работы стволовых клеток. В этом случае стволовые клетки определенной ткани при введении их в организм, находят зону повреждения и фиксируются там, исполняя утраченную функцию. Очевидно, это связано с наличием у них ключей только от своих комнат и работают только на свои типы тканей.
Преимущества применения тотипотентных веществ или стволовых клеток в лечении, регенерации или омоложении.
Индуцированные плюрипотентные стволовые клетки (ИПСК) – это клетки, которые способны образовываться из соматических клеток при введении им набора транскрипционных факторов. Именно их и смог идентифицировать Ш. Яманака, а затем перевести соматические в ЭСК-подобное состояние. ИПСК получают из соматических клеток, репрограммируя их до плюрипотентного состояния с помощью набора определенных транскрипционных факторов. Был проведен анализ влияния 24 факторов, тем или иным образом вовлеченных в процессы становления и (или) поддержания плюрипотентности.
Используя ретровирусную трансфекцию, авторы произвели трансформацию мышиных эмбриональных фибробластов различными комбинациями данных факторов. Показано, что для индукции плюрипотентного состояния в мышиных эмбриональных фибробластах достаточно использование комбинации факторов Oct-4, Sox2, c-Myc и Klf4. С практических целей ИПСК наиболее перспективны в регенеративной медицине чем ЭСК, так как здесь не нужны будут человеческие зародыши и экстракорпоральное оплодотворение. ИПСК, как и ЭСК, имеют нормальный кариотип, экспрессируют маркерные гены плюрипотентности. ИПСК имеют преимущество, в том, что могут быть получены из клеток любого пациента, а это снимает проблемы иммунологической совместимости.
Что лучше: ИПСК или ЭСК? Ответ пока наука не дает, так как тут могут вмешиваться и эпигенетические отличия, а это в последующем может проявиться отклонениями на этапе дифференцировки клеток.
Для индуцированных плюрипотентных стволовых клеток мыши в качестве основного теста на способность к дифференцировке проводят тестирование на формирование жизнеспособных химер, например путем инъекции диплоидных ИПСК в тетраплоидную бластоцисту, когда была получена мышь, организм которой полностью развился из ИПСК, что указывает на эквивалентность некоторых линий ИПСК по своей способности к дифференцировке эмбриональным стволовым клеткам.
В практических целях преимущества будут при применении в чистом виде тотипотентных веществ. Но, с другой стороны, эти вещества только начинают идентифицировать. Подходы, основанные на стволовых клетках, тоже имеют свои преимущества, но тут может проявится и побочное действие. Поскольку с нашей точки зрения применение тотипотентных веществ проще, то возможные эти методики в этой книге мы осветим более подробно.
ЖИВОЕ ИНИЦИИРУЕТ ЖИВОЕ, ЗАПУСКАЕТ МЕХАНИЗМЫ ЗДОРОВЬЯ.
Ростки растений в качестве лучшего источника тотипотентных веществ-индукторов жизненной силы = витаукт.
Только в состоянии активного роста наиболее полезны продукты как растительного, так и животного происхождения.
Живое инициирует живое. В пище должны преобладать активированные к жизни продукты, а не находящиеся в состоянии покоя высушенные семена, которые насыщены ингибиторами, а также не мёртвые, термообработанные кашки и тем более не мясо давно умерщвлённых животных. Только так можно рассчитывать на инициацию процессов ревитализации = усиление Жизненной Силы = ВИТАУКТ, ювенилизации = омоложение, клеточной репарации, регенерации и восстановления.
Нечто подобное происходит в поврежденной эндотелиальной прослойке сосуда, ведущей к атеросклерозу, так и при многих других случаях необходимости регенерации, например при циррозе печени, диабете, нефрозе, амиотрофическом склерозе, ожоговом некрозе, старческой катаракте и многих других. Тотипотентные белки совместно с мультипотентными запустят экспрессию генов до такой степени, что восстановление = оживление высоко функциональных клеток будет преобладать над процессами дегенеративными и сенелита = старения.
В пище должны превалировать живые компоненты. Здоровый метаболизм зависит от того какой рычаг уравновешивания перевесит: рычаг витализации или рычаг оксидации (окислительного стресса). Продукты распада мёртвой пищи — это индукторы давящие на рычаги усиления механизмов и программ клеточной смерти, гликации, старения, а в малых дозах – нахождения в хроническом провоспалительном состоянии, повышенной готовности, предрасположенности к болезням, а проще слабой Жизненной Силы = ВИТАУКТ.
Продукты лежалые содержат ингибиторы или тормозят метаболизм, делают его вялым, «коптящим», как бы «заливают водой топку». Особенность этих продуктов в том, что в них нет живых ферментов, способных вести к самоферментации, а вернее к аутолизу, самопереваривания за счёт собственных ферментов. Только такие продукты способные к аутолизу дадут наиболее полноценные продукты расщепления на ингредиенты, полностью подходящие нам, а не полутоксичные продукты распада. Если нет аутолиза, то разложение продукта осуществляется микробиотой нашего кишечника, экоценотический состав которой в большинстве случаев нарушен и процесс идёт по пути гнилостной или бродильной ферментации, а в меньшей степени ферментами ЖКТ. В норме 80% пищеварения должно осуществляться в кишечнике путем аутолиза.
Наша цель уйти максимально от негативной стороны действия, которая гасит жизненную силу, но при этом усиливать противоположное крыло, инициирующее жизненные начала. Наиболее ценны не просто живые, а лучше находящиеся в активной фазе распускания и роста. Это фактически самый расцвет жизни, самый её накал. Именно величина этого накала определяет жизненную силу растения, задаёт силу стимула и тембр её звучания на всю дальнейшую жизнь. От состояния этого апекса зависит будет ли это растение в последующем здоровым и мощным или будет чахнущим и слабым. Это так называемые апексы растений – их верхушечная ростовая часть. Именно эти «фитили» обладают максимальной пробуждающей жизненной силой, способной зажечь мощное пламя жизненной горелки растению вплоть на многие годы или столетия. В культуре ткани они способны расти неограниченно. Только в апексах находятся полноценные тотипотентные вещества индукторы, способные инициировать рост неограниченно. В отличие от эмбрионов животных, которые на стадии эмбриональной диапаузы теряют возможность вырабатывать тотипотентные белки, апексы растений всегда могут вырабатывать их.
Апексы всегда находятся в полярном состоянии с нижележащими частями ростка, в котором правят иные силы, то есть торможения и дифференциации. Они в наших целях не нужны. Именно эту апикальную часть растения и нужно использовать максимально в оздоровительной практике. Всё остальное как стебель, семядоли и корешки не представляют ценности. Их лучше отбрасывать.
Биогенные продукты.
Такие биогенные продукты из ростков радикально отличаются от продуктов из семян, которые являются продуктами покоя. Особенность ростков – присутствие в них живых ферментов. Основное условие здоровья кишечника, необходимого для аутолиза и подавления гнилостных и бродильных процессов и инициирования жизненных начал в организме.

На фото: Капуста брокколи – источник ростовых гормонов и полного комплекса веществ нужных для здоровья.

На фото: Капуста Романеско – лучший источник неограниченного изобилия меристемных почек, кладезя ювенильных ростовых гормонов и нутриентов для активирования жизненной силы клеток.
Капуста содержит до 6% белков и является рекордсменом по этим показателя, широкий набор аминокислот, а главное антисклеротические вещества холин, метионин, изобилие органических кислот. Всё это вводит их в ранг наиболее ценных продуктов в диете.

На фото: Ростки спаржи.
Верхняя часть побега = апекс, содержит гормоны индукторы – наиболее ценная часть, тогда как в нижней части накапливаются ингибиторы.

На фото: Молодые побеги сарсапарели – кладезь ростовых веществ и стимуляторов, обладают максимальной пробуждающей жизненной силой

На фото: Молодые побеги хвоща - весенний деликатес.
Спороносные пестики на сочных побегах. Они у многих народов считаются деликатесом. Богаты полисахаридами, витаминами С и каротином. Употребляют в пищу эти колоски до распускания зелени, пока они не накопили горечь. Можно есть и в сыром виде, замораживать, но есть множество рецептов приготовления блюд из хвоща.

На фото: Молодые побеги винограда.
Взаимосвязь стволовых клеток растений и организмов животных.
В принципе эта методика в науке уже рассматривается давно называется фитоэмбриотерапия. Дело в том, что стволовые клетки растений — это настоящие фабрики по производству рибонуклеиновых кислот (РНК), которые присущи ювенильным тканям. Исключительно важно то, что основа благотворного действия растительных меристем — это детоксикация, которую они запускают на молекулярно- клеточном уровне. Исследования, которые проводили на дрожжах, мухах и мышах, показали, что возраст пищи из родственных организмов влияет на продолжительность жизни. И на всех трех группах животных было показано, что «молодая» диета продлевает жизнь, а «старая» диета сокращает жизнь по сравнению с «молодой».
Механизм такого действия не определен, но можно предполагать, что это связано с накоплением «старческих» веществ. Молодые нежные не развернувшиеся листочки имеют меньшее количество грубой клетчатки, но при этом именно в апексах содержится максимальное количество гормонов роста, ауксина, цитокинина и других стимуляторов. Именно здесь инициируется рост побега и его жизненная сила, максимально открыты гены молодости. Естественно, попадая в организм человека они лучше будут будить его жизненную силу. Например, максимальная концентрация сульфорафана находится именно в молодых побегах брокколи. Молодые побеги особо обогащены фитонцидными эфирными соединениями. Предпочтение при выборе любого типа пищи следует отдавать молодым активно растущим экземплярам.
Стареющие и не растущие особи прекращают производство нуклеотидных веществ ДНК и РНК.
Можно предполагать, что употребление тканей растений или животных, в которых мощно запущен механизм производства нуклеотидов будет будить эти процессы и в организме человека. Это повысит многократно его защитные силы, устойчивость к агрессивным факторам среды, ускорит метаболизм, улучшит кислородный тип энергетики митохондрий, воспрепятствует анаэробной и гликолизной энергетике. Торможение механизмов воспроизводства нуклеотидов первый этап старения клеток. В культурах ткани клетки останавливают свой жизненный цикл благодаря постепенной утраты к делениям. Найдено, что стареющие клетки часто имеют два ядра и не синтезируют ДНК. Возможно, это связано с нарастанием эпигенетической гликацией = сшиванием гистоновых белков с глюкозой в рубашке генома, что ведет к неотзывчивости ядра на сигналы, а это очевидно и способствует образованию двух ядер.
Возможное побочное действие от корневой части проростков из семян.
Дело в том, что некоторые проросшие семена в сыром виде могут проявлять отравляющий эффект. Особо это относится к корешкам из проростков семян тыквы. Вкус у таких корешков и проростков крайне горький. Мною лично из таких проростков была сделана паста на блендере, в которую добавили яблоки и киви. На вкус неприятно, а к вечеру разыгрались тошнота, понос и рвота, которые продолжались всю ночь. К утру давление крови упало с 155 до 137 ед. а нижнее с 90 до 124 ед., пульс с 55 поднялся до 104. Пожелтели склеры глаз, что связано со сбоем печени, несколько дней слабость, судороги ног ночью.
Оказывается, корешки накапливают некое ядовитое вещество, которое очевидно борется с хищниками, которые могут ими питаться. Не зря семена тыквы используют при очистке от глистов. В них содержится кукурбитин, который и обладает противоглистным действием. Очевидно, очень много этого яда скапливается в пророщенных корешках. Жареные семечки такое действие не проявляют.

На фото: проростки с корешками из семян тыквы.
Молодые недоразвитые побеги растений действуют аналогично проросткам семян.
Они представляют собой образовательные ткани, или меристематические ткани, состоящих из интенсивно делящихся и сохраняющих особо высокую физиологическую активность. При этом они не только сами находятся в высокой степени активности, но и активируют все остальные нижележащие и соподчиненные с ними ткани. Они подобны ювенильным или эмбриональным тканям. Они-то и создают эффект омоложения всему растению, в котором они находятся. Точно такую же Жизненную Силу они будут передавать и организму человека. Только на фоне таких мощных биоэлектрозарядов можно максимально успешно воздействовать на застарелые атеросклеротические перерождения и повернуть процесс на путь регенерации.
Напомню, что эффект омоложения в случае с пересадкой печени начал проявляться на второй и третий год. Следовательно, такая диета живыми ростками и зародышами, очевидно, проявит себя при длительном применении. Гарбузовым Г. А. предложено к указанным всем методикам подключить и диету с преобладанием в ней в первые месяцы живых зеленых коктейлей из активно растущих ростков! Перед употреблением они должны быть напитаны светом – только тогда их хлоропластовые аккумуляторы будут заряжены полностью. Максимально высокий с минус-зарядом ОВП через желудок и кишечник передают организму только зеленые хлоропласты!
Гормоны растений влияющие как факторы роста и омоложение на организм животных.
Цитокинины (производные аденозина) - гормон молодости растений, сдерживания клеточного старения, который долгое время считали, что они присущи только для растений.

На фото: представлены листья махорки, половину которых обработали цитокинином, что привело к остановке старения листа в виде пожелтения.
В последние годы все же найдены цитокинины и в животных тканях (правда, пока только в культуре фибробластов). Удивительно то, что и здесь они выполняют тужу функцию сдерживания старения. Механизм действия не известен, но имеются данные что один из цитокининов – кинетин – препятствует возникновению дефектов в рибонуклеиновых кислотах при их «созревании» (сплайсинге).
А ведь фибробласты – это зародыши соединительной ткани, которые участвуют во многих функциях, в том числе отвечают за регенерацию. Напомню, что регенерация во взрослом состоянии организма животных в большинстве видов тканей крайне ограничена. Кроме того, любая ткань периодически должна самообновляться. Это означает, что стареющие и изнашивающиеся клоны клеток должны уходить, а на их место должна приходить молодая замена. В большинстве видов тканей этот процесс крайне замедлен или исчерпан. Взять, к примеру восстановление пигмента меланина в луковицах волос, который связан с выработкой стволовыми клетками этого пигмента. С возрастом стволовые клетки теряют свою активность и это приводит к седине волос – четкий признак старения, который несколько подобен пожелтению листьев растений, когда в них исчезает пигмент хлорофилл. Но на фото четко видно, что с помощью гормонов можно остановить пожелтение листа = его старения.
Почему стволовые клетки теряют с возрастом активность, а волосы седеют? Очевидно, это связано с генетическими возрастными программами на отключение. Но определенную высокую роль здесь, очевидно, играет и изменение гормонального профиля. У растений такая закономерность здоровья листьев четко прослеживается в зависимости от места расположения листа в кроне и ослабления апикального доминирования, когда уровень гормонов в нижних частях кроны резко уменьшается. В организме человека на ограничение роли апикального доминирования могут играть накопление клонов стареющих клеток.
Следовательно, в организме человека тоже можно поддерживать нечто подобное «принципу поддержки апикального доминирования», то есть стволовых клеток. Этот принцип хорошо просматривается в эпифизах костей, ростовых точек ногтей, волос. В коже правят фибробласты и в которой особо четко просматривается признаки старения кожи в виде её увядания, что связано с ослаблением репаративных процессов ней. Механизм аналогичный пожелтению листа. Только в растении старение включается в каждом листе как благодаря локальным особенностям гормонального фона, так и благодаря сезонным гормональным перестройкам. В организме человека аналогом сезонного старения являются общеорганизменные возрастные гормональные перестройки, например климактерические. Гормональный фон первоначально меняется не на уровне клеток, а на внешних уровнях, то есть на уровне всего организма или ветвей в кроне, или тканей.
У растений апикальные меристемы в принципе потенциально могут давать бесконечное число клеточных делений и перезакладки новых почек.
В животных организмах эти возможности ограничены эффектом Хейфлика, то есть в культурах ткани наблюдается не более 60 кратное деление фибробластов. До сих пор считают, что этот эффект связан с укорочением теломер на хромосомах, которые якобы и являются ключом к этому ограничению. Но тем не менее клетки человека взятые на стадии зиготы могут проявлять бесконечное деление. Следовательно, ограничения включаются за фазой зиготы и не являются в полном смысле запрограммированы изначально, генетически, а включаются на определенном этапе эмбриогенеза или взрослой особи. Нечто подобное топофизису у растений, когда клоны побегов многократно раз повторяют свою особенность роста в кроне, например плагиотропность или закладывать репродуктивные, а не вегетативные почки. Эту память они несут уже в самих в себе, а не получают указания с внешних уровней регулировки организма.
Если выдвигаемые мною предположения правильны, то эффект Хейфлика можно обойти, если для этого использовать белковые тотипотентные ключи, которые работают еще в зиготе. У животных эти ключи специфичны и не подходят для стадий развития после зиготы, тогда как у растений эти ключи более универсальны и позволяют растению осуществлять неограниченное количество митозов.
ВОЗМОЖНОСТИ РЕЮВЕНИЛИЗАЦИИ = ОМОЛОЖЕНИЯ ОРГАНИЗМА С ПОМОЩЬЮ ИСПОЛЬЗОВАНИЯ РАЗЛИЧНЫХ ПРИРОДНЫХ СТИМУЛЯТОРОВ ЖИВОТНЫХ, НАСЕКОМЫХ И РЫБ.
Ювенильные гормоны насекомых в качестве лучших источников тотипотентных веществ-индукторов молодости = жизненной силы = витаукт.
Они в первую очередь тормозят метаморфоз, но при этом являются индукторами роста и развития личинки, не дают преждевременно перейти в метаморфоз в куколку, а затем в имаго. Ювенильным гормонам противостоят гормоны зрелости = ограничители и общий результат перехода в новый этап развития зависит от соотношения этих противоположных гормонов. Таким образом гормоны ограничители не всегда могут полностью выключать гормоны ювенильности, в этом случае происходит количественная регулировка, а не односторонняя одним гормоном.
В человеческом организме, очевидно, происходит нечто похожее, когда слабеет уровень гормонов (ювенилизации) и усиливается уровень гормонов ограничителей? Во взрослом организме все же могут появиться тотипотентные и плюрипотентные белки, например в плацентной жидкости у матери и это не мешает реализации онтогенетического развития. Фактор количественного соотношения ювенильности и ограничителей здесь срабатывает в пользу последних. Общая программа онтогенеза здесь не отменяется из-за небольшого присутствия ювенильных начал. Для ювенилизации нужно более весомое крыло этого маятника.
Личинки насекомых.
Данных о их лечебных и пищевых свойствах множество. Например, личинки восковой моли имеют рекомендации в качестве противовирусных и кардиопротекторных. Интересны данные по применению их при раке, где способствует улучшению качества жизни больного человека.
Личинки пчел в оздоровлении и лечении.
В их жировом тельце находится половина тех же веществ, которые есть в маточном молочке. Однако некоторые исследования свидетельствуют о том, что целебные свойства личинки пчелы превосходят молочко. В личинках собран большой запас разных групп ферментов и гормонов. Благодаря этому лечение личинками пчел показано при большом количестве заболеваний. Средство эффективно при стрессах, для лечения щитовидной железы и восстановления мужских половых функций. Также его используют для восстановления давления, ведь оно способствует расширению сосудов.
Маточное молочко пчел – «королевское желе».
Напомню, что обычная пчела живет 56 дней, тогда как пчелиная матка живёт многократно раз больше, что связано с питанием её не мёдом, пыльцой и нектаром, а маточным молочком, которое содержит фолиевую кислоту (В9). Оказалось, что именно она (не белки и РНК) селективно отвечает за такой путь развития личинки в матку.

На фото: личинки пчел в сотах.

На фото: личинки муравьев.
Плацентарная жидкость – экстракт плаценты.
Применение плацентарных экстрактов относится ближе к лечебным методам чем к реювенилизации. Уточню, что экстракт плаценты это всё же набор фетальных белков (поздних этапов эмбриогенеза, стимулирующих готовые ткани), а не тотипотентных белков, стимулирующих универсальную регенерацию.
Любое лечение предусматривает временное воздействие, тогда как оздоровительные методики подразумевают практически постоянное применение. Но, очевидно, важна не столько эта сторона химических трансформаций и биологических действий веществ из них, а феномен митогенетического воздействия, электрохимического сигнала, приводящего к активации жизнедеятельности клеток и ткани, в том числе усиливает митозы.
Показано оптическое взаимодействие дробящихся яйцеклеток лягушки, проявляющееся в стимуляции «передовыми» яйцеклетками отстающих «соседей».
Ювенильные гормоны из икры земноводных в качестве лучших источников тотипотентных веществ-индукторов молодости = жизненной силы = витаукт.

Ювенильные гормоны из икры и мальков рыб в качестве лучших источников тотипотентных веществ-индукторов молодости = жизненной силы = витаукт.
Зародыши рыб помогут искусственно получать стволовые клетки.
В таком направлении работают ученые по изучению возможности использования стволовых клеток от эмбрионов на примере рыб данио-рерио. Для этого ведутся работы для отработки биотехнологий для искусственно созданных условий для выращивания стволовых клеток in vitro. Эти технологии и клетки особо перспективны в медицине для лечения ряда неизлечимых заболеваний.
Очевидно, еще более перспективно разрабатывать технологии не просто размножения стволовых клеток рыб, но и получения из них тотипотентных веществ. Считается что эти тотипотентные и плюрипотентные вещества-индукторы одинаковы у рыб, птиц, земноводных и человека.
Репликативный гомеостат = репликастат и репликарез – механизмы, определяющие реализацию онтогенеза.
Описанные выше тотипотентные и плюрипотентные вещества-индукторы можно считать инициаторами точек роста и поддержкой их непрерывной молодости. Если бы на них не накладывалось ингибиторное регулирующее действие клеток ограничителей, то в животном мире в принципе возможна непрерывная молодость и рост, как например у гидр. У гидр, как известно, процессы появления новых клеток уравновешены с процессом отмирания старых и так до бесконечности.
Репликастат - это так называемый ростовой репликативный гомеостат, отвечающий за динамическое равновесие роста и отмирания. У всех остальных животных этот репликастат перекошен на разных этапах онтогенеза в ту или иную сторону, что и определяет особенности их возрастных ограничений.
Репликарез – это противоположно действующий принцип регулировки механизмов неуравновешенных репликаций. В какую сторону будет работать репликастат записано в гено-программах в каждом этапе онтогенеза. Термины репликастат и репликарез впервые предложены Гарбузов Г. А.

На фото: рыбки Данио Рериа стволовые клетки которой изучают на предмет использования регенерации эндотелия у человека.
Могут ли приёмы повышения электропотенциала абсолютного здоровья содействовать реювенилизации организма?
Выше мы говорили, что заряд -200 mv соответствует абсолютному идеалу здоровья, когда возможен максимальный потенциал защитных, регенеративных и репарационных сил, способных легко смести последствия или противостоять практически любой патогенной нагрузке, приводящей к огромному числу хронических «неизлечимых» «Болезней Цивилизации». Он является индуктором Жизненной Силы = ВИТАУКТ, показателем свежести, расцвета всей мощи здоровья организма. Задачей является пролонгирование его сквозь всю жизнь, даже вопреки внутренним механизмам развития и старения.
В то же время, все вышеописанные приёмы по пробуждению жизненной силы с помощью живых продуктов как проростки, зародыши и тому подобное явно будут не достаточны. Скорее всего, они дадут небольшой положительный эффект, но это явно не радикальные методики. Онтогенетические программы развития и груз жизни всегда будут превалировать. Задача запустить те механизмы, которые в норме никогда не запускаются. Мощь повышения биопотенциалов будет недостаточна. А можно ли её усилить и тем самым запустить обратный процесс, механизмы регресса «неизлечимых» болезней. Предполагаю, что это возможно достичь при особых дополнительных условиях. Те дозы живых зарядов, которые можно получить от ростков и зародышей не позволят в достаточной мере поднять жизненный потенциал. Его можно усилить, например, увеличением объёма очищенных ростков от семядолей растений. Дело в том, что основную ценность в семени представляют не семядоли с их эндоспермом, в которых хранятся все питательные вещества как жиры, белки и углеводы, а именно меристема самого ростка. Чтобы усилить их действие, можно эти ростки активно отделять от семядолей, получая более высокий концентрат действующего начала. Желательно ежедневно получать до 100 г таких апексов. Следует знать, что для этого необходимо использовать семян в десятки раз по весу больше, чуть ли не 1-2 кг или более ежедневно. Тем не менее, и это не изменит радикально ситуацию.
Голодание – один из приёмов противодействующих старению.
Очевидно, здесь важно подключить запуск и других реювенилизирующих механизмов. Лучшим из них будет лечебное голодание, вернее принимать эту активирующую организм биомассу на фоне голодания. Это запустит одновременно механизмы клеточной аутофагии и поведёт к выбраковке ослабленных, сенелитирующих клонов клеток, мешающих запуску здоровых клеток и тем самым запустить не только механизмы репарации, то есть на клеточном уровне, но и механизмы регенерации, то есть на уровне всего организма.
В книге мы обсуждали, что голодание – лучший способ противодействовать воспалению, что доказано научно. Подавляя механизмы воспаления, мы усиливаем механизмы противодействия, репарации, восстановления, которые начинают превалировать над патологическими рычагами. Но подавить голоданием провоспалительные процессы в принципе дело тонкое. Просто обычный длительный голод, наоборот, запускает механизмы стресса и ослабляет организм, запускает механизмы воспаления. На фоне голода приёмы стимулирования жизненной силы энергией ростков создадут конфликтную ситуацию двух противоположных процессов, в которой одна из сторон пытается повести на путь эмбрионализации, неотенирования. Всё зависит от того какая сторона конфликта перевесит и создадутся особые условия «межвластия генов», неподконтрольности, когда могут запуститься, проснуться древние заглушенные механизмы регенерации, например, как у аксолотлей.
Голодание при этом глушит одни программы, например роста, пролиферации, а ферменты и белки-стимуляторы ростков будет запускать, потенцировать противоположные программы, которые присущи ювенильным тканям. В этом случае на фоне снижения простого роста будут активизированы более тонкие пластические и репаративные функции клеток. Таким образом, будет происходить перепрограммирование. Такое сочетание двух противоположно направленных приёмов по индуцированию и оживлению тканей на фоне торможения их роста из-за отсутствия достаточного количества пластических (питательных) веществ должно привести к запуску обратных механизмов (как мы полагаем), а именно регенеративных. Это запускает те генетические программы, которые в обычной жизни плотно заглушены и организм ими не может никогда воспользоваться. Голод открывает наглухо закрытые двери. Этому содействует и аутолиз. Предлагаемое количество чистых ростков 100 г в сутки будет по калорийности аналогично длительному голоданию, но по комплексу живительных процессов будет аналогично тем процессам, что происходят в юности.
ВОЗМОЖНОСТИ РЕЮВЕНИЛИЗАЦИИ = ОМОЛОЖЕНИЯ ОРГАНИЗМА С ПОМОЩЬЮ ИСПОЛЬЗОВАНИЯ ЗАРОДЫШЕЙ ЖИВОТНЫХ С ЦЕЛЬЮ ПЕРЕКЛЮЧЕНИЯ ОНТОГЕНЕТИЧЕСКИХ ПРОГРАММ.
Особую важность представляет применение живых яиц птиц.
Однако намного лучше применять пробужденные, то есть подвергнутые инициации в инкубаторе 2-4 дня. В таком виде в них запускается в начале производство тотипотентных (универсальные генные пускатели), а затем фетально-эмбриональных белков, последние из которых отвечают за экспрессию = включение генов и включение, и последовательное разворачивание программ на более конкретную специализацию органов и тканей. Все они являются индукторами запуска программ на восстановление, регенерацию, омоложение. Таким образом, создаются зачатки определённых линий стволовых клеток, которые в последующем получат свою специализацию. Таким образом, в яйцах, как и в апексах растений, самым ценным является зародыш, а не его желток и белок. Здесь в зародышах находится весь набор ключей.
Следует понимать, что количество тотипотентных ключей в зародыше яйца предназначено только для определенного небольшого количества клеток зиготы. Следовательно, то количество этих белковых ключей, находящихся в желтке, будет крайне недостаточно для организма человека, размеры которого в тысячи раз больше. Понятно, что для наших целей нужно будет использовать множество желтковых зародышей в течение длительного периода.
Также следует понимать, что пробуждающий эффект тотипотентных ключей будет нивелироваться действие стареющих уже имеющихся клонов окружающих клеток. Как говорится: как откроется, так и закроется. Действие ключей экспрессоров должно преобладать над действием ключей репрессоров. Их сигнал должен быть сильнее сигнала второго. Концентрация тотипотентных белков должна быть выше намного репрессирующих начал. Учитывая, что на разных этапах онтогенеза подключаются разные гено-ключи, считаю, что для этих целей следует воспользоваться максимальным набором ключей. Это означает, что надо будет использовать зародыши яиц одновременно разных ступеней пробуждения, то есть в один день использовать зародыши и 2 – 3 – 4 – 5-дневного развития.
Подытожу, наиболее ценными являются пробужденные яйца на фазе, в которой преобладают тотипотентные белки, отвечающие за закладку и инициацию первичных стволовых клеток = мультипотентных, то есть универсальных клеток, из которых могут закладываться любые органы, а не более специализированные линии клеток под определенные ткани. Тотипотентные белки могут появляться исключительно на начальной эмбриональной паузе развития. В последующем их появление полностью заблокировано.

На фото: хорошо видно канатик у желтка, это некая пуповина, но не зародыш
На фото: Бластодиск оплодотворенного яйца – зародыш в желтке

Научные исследования и доказательства терапевтического действия некоторых ростовых факторов яйца.
В настоящее время имеется определённая научная база с первичными клиническими исследованиями, показывающих конкретные терапевтические возможности применения живого белка яиц при лечении ряда заболеваний путём применения его в виде уколов. На метод имеется патент. Способ извлечения в чистом виде генных белковых ключей пока не разработан. Известен и метод Капустина Г. А., который применял яичную массу в смеси с пятью частями физиологического раствора. 5 мл этой стерильной смеси вводили в мышечную ткань. Имеются отзывы частнопрактикующих людей, что удалось избавится от неизлечимых миомы, кисты, артроза. В тоже время, у метода применения такого яичного белка в виде уколов имеются и недостатки, в том числе и возможная аллергическая реакция из-за наличия чужеродных антигенов, вплоть до шоковой реакции Квинке, некроза ткани и др.
Кроме того, метод не подразумевает воспользоваться пробужденными яйцами, в которых инициировано производство генных ключей. Следовательно, механизм его действия принципиально отличается от того, что предложено нами. Скорее всего, суть этого метода заключается в активировании врожденного иммунитета и не связан с генетической экспрессией. Возможно, в этой яичной массе и имеются заранее заложенные тотипотентные ключи, которые просыпаются или инициируются за счет термовоздействия. Тепло может действовать как фактор разворачивания конформационных = пространственных белков. Они распаковываются и начинают действовать, экспрессируя = включая определенные генные программы.
Тем не менее, метод введения яйца уколами удивляет тоже рядом описанных феноменов излечения ряда хронических заболеваний. При этом в организм вводится около 1 мл смеси яичной массы и этого достаточно, чтобы запустить некие механизмы самовосстановления. Известна книга доктора Кривонос про опыт применения уколов из куриных яиц в лечении. Он приводит описание научного эксперимента, подтверждающего эту методологию. Эксперимент проводили на мышах, которых подвергли мощному рентгеновскому летальному облучению, при котором происходит 100% гибель животных. В группе, где вводили белково-желтковую смесь живыми остались 70%. Такой эффект связывали с уменьшением радикаловых последствий от излучения и ускоренной репарацией поврежденных ДНК.
Мною выдвинуто предположение, что это связано с наличием гено-ключей, которые способны запускать репарацию и регенерацию, то есть программ, выключенных у детерминированных клеток. Он описывает случаи излечения при нарушении иннервации стопы = повреждение периферического нерва, утверждает об улучшении памяти, улучшение работы иммунной системы за счет излечения от рожистого заболевания, восстановления гормонального фона, что проявлялось в виде избавления от мастопатии. Описан случай больной с неработающей почкой, когда после 20 инъекций удалось восстановить почку 48%. Известна работа Джона Р. Девидсона, который в 1929 г работал с экстрактом 9-ти дневного оплодотворенного куриного яйца, то есть пробужденного, и утверждает, что именно оно имеет наивысшую энергетическую силу для оздоровления. Он вводил этот экстракт смертельно больным онкологией. При этом процент выздоровления был очень высок. Очевидно, здесь уже имелись плюрипотентные белки, которые используют в качестве фактора роста фибробластов в культурах ткани. Эти же идеи реализовала для практического применения компания LifePharminc в виде капсул Лэминайна. Утверждают, что якобы она несет информацию о здоровой клетке в те органы, где произошёл сбой, и клетки начинают питаться его компонентами и восстанавливаться до первоначального состояния. Происходит омоложение клетки и оздоровление всего организма. Показывают, что Ламинин это некий белок из яйца, который присутствует в организме только в эмбриональной фазе у любого животного организма. Препарат прошел клинические испытания и сертификацию.
Основное действие связано с факторами роста фибробластов. Во взрослом организме этот фактор участвует в ангиогенезе, заживлении ран, пролиферации и дифференциации. Они есть и у взрослого организма в плаценте и женском молоке, обеспечивая восстановительные механизмы, в том числе и для поврежденных клеток и программирует стволовые клетки. В более взрослом состоянии организм их уже не вырабатывает. Интересны его показания для применения человеком. Компания, которая его выпускает утверждает, что если у вас есть проблемы со здоровьем, то результат можно увидеть только с 4-го месяца приёма, вплоть до улучшения зрения, качества волос и возврата их цвета. Следует обратить внимание, что при длительном применении следует ожидать восстановление нормального уровня кортизола = гормона стресса, способствует выработке коллагена, с которым связана старческая саркопения, то есть уменьшение и одряхление мышечной массы, улучшает память, стабилизирует давление, регулирует сахар в крови. Есть утверждения, что комплекс Laminine Omega+++ можно использовать для очищения сосудов и в таком комплексе он усиливает действие другого препарата Ламинина в 4 раза, за счет чего полностью очищает артерии, повышает эластичность сосудов, нормализует липидный обмен в организме и восстанавливает всю сердечно-сосудистую систему.
Способ применения живых яиц – источника белков генетических ключей для запуска ювенильных программ.
Неустойчивость генетических-ключей при приёме их внутрь.
Следует понимать, что в роли тотипотентных регуляторов = генетических ключей могут быть РНК и ДНК или белки – это особые, возможно конформационно-пространственные белки (работают не за счет аминокислотного набора, а за счет конфигурации в пространстве), которые мало того, что не термостойки, так еще подвергаются денатурации и уничтожению в кишечнике под действием кислот, которые имеются в желудке, а также желчи и щелочных ферментов в 12-ти Перстной кишке и Тонком кишечнике. Так попадая в ЖКТ жидкий живой яичный белок и желток, подвергаются таким же преобразованиям, денатурации аналогичной той, как и при варке их в кипятке, превращаясь в твёрдую сваренную вкрутую массу.
Способы приёма живых яиц по Гарбузову Г. А.
Для решения барьерной проблемы и доставке неповрежденных гено-ключей в организм Гарбузов Г. А. впервые предложил способы приёма содержимого яиц в виде «гоголь-моголя» из пробуждённых живых яиц перорально (через рот) или ректально с помощью клизм. Особенностью способа является приём в ранний утренний период, когда желудок, поджелудочная железа и желчный пузырь будут находиться в неактивном состоянии покоя.
Сублингвальное применение зародыша = под язык или за щеку.
Чтобы не возбуждать включения желудочного рефлекса, например, по выделению соляной кислоты в желудке, а в 12ПК и ниже ферментов и желчных кислот, на пищеварение при попадании в ЖКТ определённой пищевой массы, нами предложено использовать не всё яйцо целиком, а выделить только его зародышевую часть, смотрите фото.

На фото: способ экстирпация – выделения зародыша из желтка с помощью шприца.
Применение гоголя-моголя из смеси зародышей – один из оптимальных приёмов: для этого берут зародыши по отдельности на 2-ой, 3-ий, 4-ый и 5-ый день развития. В целом это будет меньше чайной ложки, что недостаточно для срабатывания пищеварительного рефлекса.
Экстирпация зародышей: их лучше отсасывать с помощью стерильного шприца без иглы, которые остриём максимально близко подводят к зародышу, а затем отсасывают его до 0,5 – 1 см. куб. При этом такая масса будет быстро продвигаться вдоль этих органов с нескольким чайными ложками тёплой воды, легко минуя эти преграды и попадёт в тонкий кишечник. Здесь эта масса должна легко всосаться в кровь, не подвергаясь распаду, и принося в организм эти ценнейшие регуляторы. Оптимальным временем для их приёма будет ночной период, когда организм находится в сонном состоянии. Больше всего этому соответствует ночной период около 4-6 часов.
Сублингвальный = подъязычный приём экстирпата - другой возможный вариант приёма частиц зародыша путём рассасывания его там – 5-10 мин. Поступление в кровь обеспечивается богатой васкуляризацией слизистой оболочки полости рта.
Доза-эффект. В медицине существует такое понятие, которое описывает результативность действия данного вещества в зависимости от его концентрации. Предполагаю, что количество этого вещества-ключа = тотиптентного в одном зародыше крайне недостаточно на размеры организма человека. Конечно, все задаваемые вопросы здесь пока однозначного ответа не получат, а только гипотетические. Нужны разработки технологий извлечения этих ключей, определить эффективную дозу, найти ответ что представляют собой эти вещества. Но наука часто делается энтузиастами, которые порой делают то, что смогут целые научные лаборатории. Предположительно, можно ожидать что эффект можно получить от смеси водной экстракции 10 зародышей от 2-3-4-5-9- дневных зародышей в течении 3-4 месяцев, которые вводят через 1-2 дня под кожу или внутримышечно по 2-5 мл.
Методы лечебного голодания для усиления эффективности других методик реювенилизации.
Старение на клеточном уровне во многом связано с деградацией лизосомами. В них находятся ферменты по расщеплению как пептидов, нуклеиновых кислот, полисахаридов и других загружающих цитоплазму веществ. Чем менее активны лизосомы, тем больше в них мусора, а значит и ускорение старения клеток. Голод здесь окажет определенную помощь.
НЕОТЕНИЯ = ЮВЕНИЛ В ЦАРСТВЕ РАСТЕНИЙ - ПРЕОБЛАДАНИЕ ВЕГЕТИРУЮЩИХ НАЧАЛ НАД РЕПРОДУКТИВНЫМИ.
Анализ неотении и ювенилизации на примере растения секвойи показывает, что ювенильность всегда совпадает с преобладанием вегетативного состояния. Чтобы проанализировать сущность старения у деревьев сделаем это на удобной модели: например, на хвойном дереве лиственнице.
В природе в лесу дерево начинает давать признаки старения довольно в позднем возрасте: чаще всего в 50-70 лет, что проявляется в замедлении роста прироста главного матрикального, то есть верхушечного побега, а также многих других побегов на окружающих вокруг ветвях. Из-за этого меняется форма кроны. На плагиотропных побегах появляется множество стробил = шишек = генеративных органов (мужских и женских). Чем старше дерево, тем выраженнее становится появление генеративного типа побегов и изобилие шишек на них. В этом возрасте 50 лет начинается полная зрелость = адальтус и продолжаться он может до 100 – 200 лет, а затем приходит состояние матуры = перезрелости, когда рост побегов максимально ограничивается, чуть ли не до нуля, но дерево при этом обильно плодоносит вплоть до самоистощения и самоугнетения непомерным стробилообразованием. Происходит явное угнетение вегетативной фазы, но чрезмерное преобладание, засилие генеративной. У животных это когда рост заменяется развитием. Фаза матурации (перезрелости) постепенно переходит в фазу сенелита = старения дерева. Сенелит сам не приходит, а его включает фаза матуры = перезревания.
Здесь следует обратить внимание на это утверждение, так как ниже мы будем показывать, что нечто подобный процесс происходит и в биологии онтогенеза человека. В природе для эволюции фазы онтогенеза ювенильность, адальтус можно растягивать до неограниченного количества лет, если это выгодно для существования вида. Но эти свойства должны быть записаны в генетических программах, которые их обеспечивают. Эти свойства и осуществляются при необходимости в филогенезе многих видов растений. К примеру, растения могут иметь 1-летний, 2-летний и т.д. циклы существования и вплоть до нескольких тысяч лет. Все зависит от потребностей эволюции, а ограничений принципиальных для этого нет. Никакого ограничительного эффекта Хейфлика здесь нет.
Нами понимается эффект Хейфлика, как переключение на новые программы развития, осуществленные в постзиготный период развития или эмбриональной диапаузы, когда тотипотентные программы успевают уже быть заблокированы и клетки превращаются в прогениторные. Они уже обладают жесткой детерминацией, двигаться только вперед по очередным программам с усеченной возможностью к возврату в исходное состояние. Возможность того, что они могут быть разблокированы и переведены в предшествующую программу хорошо просматривается у онкологических клеток. Чем более растянута фаза матуры (сверхзрелости), тем позже может наступить сенелит. Например, у дерева секвойи этот период матуры может растягиваться до тысячи и более лет. Есть виды деревьев способные доживать до 4 тысяч лет. Следовательно, у древесных нет такой проблемы-ограничителя жизни на клеточном уровне как эффект Хейфлика. Таким образом, сенелит как феномен в природе не является неизбежным и не лежит в сути всего живого. Повторяю, не является неизбежным, а всего лишь генетическая реализация, закрепленная в филогенезе, если бы надо было для пользы вида, то сенелит мог бы быть исключенным как фаза онтогенеза. Так, например, известна самоклонирующаяся колония креозотового куста (Larrea tridentata), возраст которой оценивается в 11700 лет.
Среди высших древесных растений сенелит регулируется не изнутри клеток, а на уровне степени развития всего организма, продвижения по ступеням очередного гормонального профиля. Каждый этап развития имеет свой гормональный профиль. Именно наступление соответствующей очередной ступени гормонального профиля запускает новые программы по переходу на новый этап онтогенеза. Это означает, что переход на новый этап онтогенеза = фазу развития осуществляется по мере его становления, степени зрелости его апикальных меристем, а значит и соответствующим ему гормональным профилем.
Сенсорами = чувствительными тканями к гормональному профилю являются апикальные меристемы. Они как дирижёры задают тон всем остальным ростовым точкам = соподчиненным меристемам и тканям. Между апикальными меристемами и соподчиненными имеется прямое и обратное взаиморегулирование, то есть влияние друг на друга. В зависимости от того, чье влияние сильнее будет зависеть особенности дальнейшего развития и существования организма. Понимание этих принципов регулировки онтогенетических фаз развития на примере древесных растений позволит легче понять аналогичные принципы регулировок в организме человека.
Вегетативное размножаемые клоны могут тоже существовать тысячи лет. Ограничениями для существования, например гигантского дерева секвойи является утрата оптимального соотношения поставки питательных веществ из корней в крону. Из-за гигантских размеров для дерева становится неоптимальным тянуть нутриенты вверх в крону. Это рано или поздно приводит к ухудшению питание ветвей в верхней части кроны, а это в свою очередь является причиной включения генетических программ на вход в состояние матуры, то есть перезревания, которое и является стартом для включения механизмов входа в этап сенелитического развития, а затем и перерождения.
Ускорение наступления фазы старения у растений.
В среднем, например, период наступления матуры или сенелита у лиственницы в 50-150 лет. Но тем не менее можно обнаружить экземпляры, у которых все признаки старения начались в 10 лет. Так наблюдения за посадками этих деревьев в городских условиях в каменистую бедную, неглубокую почву показало через 2-3 года все признаки старения и подготовки дерева к преждевременному отмиранию. Это проявлялось в почти полном отсутствии прироста: но при этом обильной закладке стробил, что говорило об ускоренном созревании растения.
Проанализируем особенности роста хвойного растения на примере лиственницы, которая находится в плохих условиях, например тяжелая каменистая почва. В этом случае можно отметить, что прирост её побегов крайне ограничен, но зато идёт ускоренное созревание и закладывание обилия стробил. В принципе это путь ускоренного старения дерева. У дерева длинная вегетация – это период его молодости.
Ниже приведены фото дерева туи восточной, крона и побеги которой проявляют все признаки старения: угнетенный искривленный центральный верхушечный побег. Все боковые ветви перегружены шишками, которые угнетают рост ветвей и дерева в целом. Генеративная перезрелость = стадия матура онтогенеза – дорога к сенелиту.

На фото: Стареющий клон дерева туи восточной. Старение проявляется в плохо обветвленной кроне, слабым угнетенным приростом побегов и особенно у главного центрального побега; ветви перегружены обилием мелких шишек, которые истощают растение. Явно выраженное преобладание генеративного развития над вегетативным, что соответствует фазе онтогенеза матуры или аналогичному процессу прогерии у животных.

На фото: Представлен побег с того дерева туя восточная, где четко выражена высокая степень стадийной зрелости в виде переизбытка заложения шишек = генеративных органов. В то же время угнетена вегетативная фаза растения. Это говорит о том, что растение находится в стадии матуры = перезрелости, а в связи с ограниченным ростом апикальных побегов и особенно матрикального = центрального побега, говорит о сенелите растения.

На фото: внешний вид туи западной в молодом возрасте.

На фото: Роща секвойи в Сочи в возрасте 60-лет, проявляющая все признаки суховершинения и преждевременного старения = сенелита = прогерия.
На фото видно 60-летние угнетенные, умирающие деревья секвойи. Причем это не простое патологическое усыхание деревьев, а им присущи все элементы старения, которое в данном случае проявляется в 25 раз ускоренно.
Ниже на фото для сравнения представлены секвойи 500 лет, но на родине.

На фото: Деревья секвойи на родине в Калифорнии в возрасте 500 – 1000 лет хорошо себя чувствуют и имеют нормально обветвленную вершину. Доживаемость таких деревьев достигает 2000 лет.
Таким образом, деревья в Сочи проявили преждевременное старение = прогерию. Следует признать, что прогерией более правильно считать не только феномен врожденного, то есть на уровне генетических поломок, ускоренного старения, как у некоторых людей, но и часто встречаемое приобретённое старение, как у людей, так и у растений.
Конечно, прогерия это усеченный вид старения, когда выломаны некоторые программы, предотвращающие или отодвигающие = противодействующие ему. Прогерию можно считать как наследственной, то есть записанной в генах, так и приобретенной, когда это связано с внешними факторами, но без записи на генах. Поэтому в чистом виде прогерия зацепляет не весь комплекс программ и процессов, ведущих к обычному старению. Больше похоже, что прогерия это элемент нарушения последовательности в какой-то морфогенетической программе, которая регулируется преимущественно на клеточном уровне, то есть определяется с нижнего этажа. Доказано, что прогерия связана с генетическим нарушением, ведущим к митохондриальным дисфункциям, то есть неполноценно выраженной энергетике клеток. Это в свою очередь обуславливает не просто низкий уровень энергетики клеток, а в первую очередь низкую активность стволовых клеток, что не позволяет полноценно раскрыться, трансформироваться и проявляется в виде некоторой недоразвитости.
Выше мы уже обсуждали, что мощь стволовых клеток животных можно сравнивать с камбиальной активностью растений. Недостаточность камбиальной активности ведёт к угнетению роста = вегетирующей фазы у растений, что является триггером для запуска генетических программ на старение. Таким образом, прогерия это с одной стороны недоразвитость, качественная непроработанность, а с другой – преждевременное старение без прохождения стадий адальтуса и матуры.
Следует отметить, что прогерия несколько подобна ограничению способности к делению клеток = митозам, которое также проявляется в эффекте Хейфлика, когда клетки спустя заданное количество митозов, теряют такую возможность и энергия жизни у них тухнет. Это так называемое старение линий = клонов на клеточном уровне, то есть проявляется в условиях как культуры ткани, так и в организме. То есть такая врожденная прогерия и эффект Хейфлика диктуются с нижнего клеточного уровня. Но при приобретенной прогерии её механизмы запускаются внешними факторами, когда происходит переход за рамки коридора = клиренса гормезиса.
Также как и при нормальном старении прогерия вызывает быстрое развитие болезней, присущих глубокой старости, – помутнение хрусталика, атрофию мышц = саркопению, атеросклероз = эпителиальную дисфункцию, изменения в костях и суставах. В отличие от мира растений, у животных онтогенетические регулировки более сильно детерминируют с нижним уровнем контролирующих программ. Поэтому прогерийные субъекты у людей не могут достичь нормальной половозрелости, они быстро умирают, не достигнув и не пройдя через ряд программ онтогенеза, они стареют снизу, где критерием является недостаточность процессов, а не избыточность, как при адальтусе с последующей переизбыточностью или перекосами по сравнению с оптимумами, как при матуре, а затем переходящие в функциональную недостаточность как при сенелите.
Естественный сенелит в свою очередь определяется взаимодействием как верхних этажей регулировок, так и нижних программ контроля реализации общеорганизменных процессов развития. Например, при прогерии очевидно не включены механизмы адальтуса = половозрелости и не задействованы механизмы патологического сенелитического провоспаления, которое подготавливает почву для многих болезней старости.
Таким образом, генетически обусловленная прогерия — это относительное понятие преждевременного аномального старения. Не будет большой ошибкой если мы этим же термином, но с уточнением «обусловленная», будем называть наведенную прогерию, связанную с несоответствием, разрывом возможности и потребности, генетических предрасположенностей и реальными условиями важными для развития. Фактором такой приобретенной, «обусловленной» прогерии может быть несоответствие условий экосреды. Так деревья в Сочи посажены на тяжелых не глубоких каменистых почвах. Генетически секвойя приспособлена к глубоким, богатым, дренированным почвам и благодаря этому обеспечивать бурный мощный рост. Дерево гигант благодаря своим потенциальным возможностям стало самым крупным растением мира – достигает 100 м в высоту. Для такого мощного роста ему нужна как высокая камбиальная активность, так и активность корней. Но чтобы достичь высокого уровня камбиальной активности должны быть активны апикальные почки. Крона и её верхушечные побеги должны пропорционально развиваться с корневой системой. Но в данном случае происходит несоответствие требований растения и его возможности к реализации. Это воспринимается генетическими программами как угнетение вегетативной фазы, а значит укорочения фазы ювенила и адальтуса, но сопряжено с ускорением программам матуры и сенелита. Как видим, для поддержания долгожительства и отодвижения стадий матуры и сенелита нужны активные камбиальные зоны, которые по понятным причинам у дерева резко недостаточны, а это прямая дорога к сенелиту.
В данном случае можно говорить о низкой пластичности, приспособляемости дерева секвойи к ухудшению почвенных условий. Это связано с нарушением активности корневых апексов. Мощь активности апексов, в которых определяет состояние верхушечных апексов в кроне. Эти два полюса апексов сопряжены. Если один полюс ослаб, то страдает и второй верхний полюс. Но такая высокая сопряженность записана не генетике. Потребности верхних апексов резко отстают от возможности нижних апексов, происходит разрыв возможностей и потребностей. Коридор такого разрыва у секвой очень низкий, что и обусловила скорую утрату ювенильности деревьями в указанной роще.
В тоже время следует отметить, что такой низкий коридор пластичности наблюдается не у всех деревьев. Например, у дерева дуб или у ели, фото которых приведены ниже, видно, что стадия сенелита у них не наступает даже в самых плохих почвенных условиях. Это объясняется особенностями их корневых апексов, даже самый малый прирост в них не ведет к наступлению сенелита в верхнем уровне апексов в кроне.
Для чего нужно это так досконально анализировать? Камбиальная ткань по сути является аналогом стволовых клеток в организме животных: чем активнее стволовые клетки, тем моложе растение. У секвойи кроме преждевременного старения рано или поздно наступает и стадия естественного сенелита, иногда через 2000 лет. Причиной этому является естественное наступление состояния диспропорций, слишком гигантских размеров, когда соотношение работы корневой системы недостаточно чтобы обеспечить потребности гигантского растения. Диспропорция корней, камбиальной активности, работы апексов и резкое самоослабление фазы вегетативной активности.
Таким образом, сенелит и ювенил у растений генетически через триггерные программы могут быть привязаны к темпам роста и камбиальной активности. У животных организмов матура и сенелит тоже привязаны генетически к активности стволовых клеток, но механизмы, определяющие активность стволовых клеток привязаны генетически или к способности или неспособности стволовых клеток к очередному делению. Такое привязывание стволовых клеток к внутренним программам клетки связано с потребностью животных организмов поддерживать постоянно ограниченный = сдержанный рост, а точнее уравновешенный с процессами самоликвидации стареющих линий клеток и замены их на новые. Здесь возможен механизм подключения «рубильника» для митозов через посредство теломеразного счетчика количества делений данного типа стволовых клеток. Регулировка онтогенеза с верхних уровней спускается на уровень клеток.
Особенности биологии молодости секвойи.
Важнейшей особенностью стадии молодости у секвойи является её чрезвычайно высокая энергия роста = Жизненная Сила = Витаукт. Благодаря высокой энергии вегетативной фазы подавлены элементы генеративной = половой фазы, дерево практически десятилетия может не плодоносить. Это так называемый пролонгированный ювенил. Такая мощь роста обеспечивает возможность дереву растягивать все остальные фазы онтогенеза до нескольких тысяч лет. Но следует отметить, что это возможно только в условиях особых экологических ниш, когда дерево только и способно задавать высокие темпы роста. Для этого нужны не только климат, но и глубокие наносные с гор почвы. Без этого дерево не может проявить весь свой жизненный потенциал, любые ограничения роста включают механизмы ускоренного адальтуса, матуры, а затем и сенелита.
Ингибирование = отодвижение программ старения у секвойи и ее гигантизм, как и у других гигантских растений, обусловлены не только высокими темпами роста, но и реализацией генеморфологических возможностей непрерывно поддерживать этот рост. Для этого необходимо образование особо высоких уровней гормонов ауксинов, а высокий уровень которых неминуемо отодвигают как матуру, так и сенелит.
Гетерозис - условие, которое обеспечило дереву не только гигантизм, но и долгожительство. Гетерозисный отбор, приспособление произошло именно по этим факторам. Как известно, гетерозисные растения узко приспособлены к особым важным для них условиям среды. Ареал существования у них крайне ограничен. Вне которых они не могут нормально развиваться, проявлять свою мощь и существовать. Гетерозис всегда снижает степень пластичности, приспособляемости к новым условиям среды.
В тоже время, многие другие виды растений способны сохранять свое долгожительство и вне подходящих для этого особых, например богатых почв, условиях. Это так называемые растения космополиты. Описание таковых примеров у дуба и ели приведено ниже. Их более высокая пластичность, приспособляемость связана с отсутствием гетерозиса.
Ауксины вырабатываются не только в кроне, но и в корнях. Бедные условия среды, экониши ограничивают возможность роста корней обуславливает недовыработку ими ауксинов, а значит сдерживают потенциал их роста. Это в свою очередь ведет к диспропорции потенциала роста корней и кроны, что в первую очередь сказывается на торможении роста вначале главного матрикального побега, а затем и постепенному отмиранию верхушечных ветвей.
Ауксины обеспечивают активность и рост меристем корней = их стволовых клеток, находящихся в корневых апексах. Таким образом, мощь стволовых клеток, апексов у корней поддерживается как за счет собственного потенциала, высокой способности нарабатывать ауксины, так и за счет стимулов извне и притока ассимилянтов. Порой основной ствол спилили, а за счет корней идет мощный рост поросли, который инициируют корни своими силами и резервами. Таким образом, дереву не обязателен стимул из верхнего апекса, если у дерева есть спящие почки, то возможна регенерация всего дерева. Но не у всех хвойных закладываются спящие почки, например, у сосны их нет и нет возможности к регенерации.
Таким образом, стволовые клетки корней тоже способны, как и верхние почки вырабатывать ауксин.
Сила роста апексов у корней регулируется не только извне, но и задана им изначально, что и определяет силу и мощь роста всего растения. Получается, что это заранее заданный параметр. В свою очередь это означает, что силу роста и долгожительство растения в данном случае определяет генетическая предрасположенность к этому.
Здесь мы опять возвращаемся к вопросу откуда берут начала механизмы старения: на верхнем уровне регулировок, то есть по мере прохождения онтогенетических стадий или с уровня клеток, возможностей или заданности их стволовых клеток? Из приведенных данных по корням становится очевидным, что долгожительство растению задают меристемы корней, а верхние апикальные почки в том числе и матрикальный побег реализуют возможности корней.
Из этих рассуждений следует, что растения с 1-летним циклом онтогенеза имеют заранее заданные параметры, обусловленные их стволовыми клетками корней? Или может быть их программируют верхние апексы, которые определяют степень зрелости, онтогенетическую стадийность? Очевидно, здесь идет двойственная регулировка онтогенеза: с верхних апексов идет сигнал о достижении зрелости, а это ведет к отключению программ на рост нижних корневых апексов.
Таким образом, долгожительство секвойи обеспечено особой возможностью образования гипервысокого уровня ауксинов. На бедных почвах секвойя теряет такую возможность дополнительной выработки ауксинов, что превращает ее в короткоживущую из-за диспропорций возможностей корней и потребностей кроны.
В отличие от секвойи, многие другие деревья, например дубы или ели (смотрите фото этих растений, растущих на каменистых бедных почвах), легче переносят недостаток ауксинов, что обусловлено генетической сниженной потребностью и это позволяет таким долгоживущим растениям сохранять живучесть и на плохих почвах, и при ограниченных темпах роста. Угнетение матриакального побега у секвойи ведет к ослаблению и усыханию роста других верхушечных ветвей.
Особенности биологии молодости у других хвойных растений долгожителей.
Не фиксированный сенелит. В тоже время высокие темпы роста и мощная жизненная сила не обязательное условие для длительности онтогенеза. Иногда более важное значение имеет нефиксированный сенелит, как это имеется например у 1-, 2-летних и других короткоживущих растений. Известны растения, которые могут расти на плохих каменистых почвах и вытягивать свою продолжительность жизни до 5000 лет. Таким представителем является сосна остистая.

На фото: Сосна остистая, возраст которой насчитывается 4850 лет.
В отличие от секвойи её ареал обитания или экологическая ниша крайне неблагоприятна, бедна для быстрого роста. Признаками ювенильности эта сосна не обладает, но, тем не менее, у неё не менее сильна Жизненная Сила = регенеративные способности, обеспечивающие ей возможность постоянно восстанавливать новые побеги взамен непрерывно погибающих старых. Особенность дерева, в том, что оно обладает высокой степенью самовосстановления, что и обеспечило ему высочайшую живучесть = выживаемость. Новые побеги непрерывно отрастают преимущественно не из апикальных, а из пазушных почек = брахиальных. Этот специфический путь восстановления и вегетирования можно сравнить с некоторой степенью неотении, карликовых форм у ели, криптомерии и у многих других хвойных. Апикальная почка с доминирующим ростом у них практически заглушена, но зато отрастают активно пазушные побеги, что придает растению густообветвленную форму с укороченными побегами.
Один из примеров этого у ели мы привели на фото в этой книге. Такая особенность брахиального типа регенеративности позволяет взрослому зрелому дереву закладывать несколько более молодые побеги, стадийно менее зрелые, чем у образующихся из апикальных = верхушечных почек. Это позволяет дереву «сбрасывать» программы зрелости, которые у них инициируются и обеспечиваются в апикальных почках с особым высоким гормональным профилем, возвращаться в предшествующую стадию развития, а значит несколько омолодится и не запускает сразу наступление фаз адальтуса и матуры.
В декоративном садоводстве можно встретить клоны таких форм у хвойных, которые называются ювенильными формами. У них отсутствует механизм апикального доминирования, что приводит к равномерному развитию как верхушечных, так и боковых брахиальных побегов, которые все вместе укорочены и стадийно равноценны. Дерево не перегружено шишками и поэтому нет чрезмерной зрелости, а значит и ускоренного старения. Такие растения имеют намного более высокий регенеративный потенциал и легко размножаются укоренением черенков.

На фото: Вид с близи густо обветвлённого побега ели обыкновенной, у которой нарушен механизм апикального доминирования и отрастание новых побегов происходит равномерно и из брахиальных почек, что сохраняет растению его ювенильные признаки и продление стадии ювенильности = неотении на многие годы. Регулировка фенотипа и онтостадии идёт первично изначально изнутри на эпигенетическом уровне.

На фото: Внешний вид 1- и 2-летних сеянцев ели имеющих полное морфологическое сходство внешности с неотеническим клоном.
На следующий год эти сеянцы утратят свои ювенильные черты, тогда как у клона эта ювенильность сохранится навсегда.

На фото: Приведен пример противоположный предыдущему, когда у клонового привитого растения, наоборот, преобладает апикальное доминирование, но крайне недоразвито брахиальное отрастание побегов. Это привело к отклонению от нормы форм ветвления с преобладанием только плагиотропных и апикальных побегов. На дереве присутствуют шишки, что говорит о стадийной зрелости побегов. Регулировку состояния фенотипа и онтостадии определяют сугубо внутренние факторы, а точнее на эпигенетическом уроне.

На фото: Растение, найденное в природе с нетипичным ветвлением, когда отсутствуют боковые побеги и рост осуществляется только из апикальных почек.
В природе такая особенность развития дерева связана с чрезмерной апикальностью, которая повышает степень онтостадийности, что ведёт к ускоренной крайней матуре, а затем к быстрому сенелиту и гибели. Своеобразная прогерия у растений – противоположность неотении.
Неотения и прогерия являются одними из рычагов эволюции.
Такая прогерия осуществляется за счет наращивания и достижения крайней степени элевационного = «конвейрного» эффекта, когда одна стадия ступенчато подводит к трансформации в другую; при этом гормональный профиль не совпадает с оптимумом для жизнедеятельности особи и стимулирует осложнения в сенелитической форме.
Состояние верхушечных апексов и степень их онтостадийности может определять состояние полюсно противоположных апексов корней.
Недостаточность одного из полюсных апексов ускоряет онтостадийность = элевационный механизм зрелости, перезрелости и старости. Только благодаря перепрививке прогерийного клона на более молодой подвой это придаёт клоновому привою баланс между двумя полюсами, то есть верхними и нижними (корневыми) апексами, что = жизненной силе и сохранности. В противном случае растение бы вышло за рамки гормезиса. Сдерживание этого обязано благодаря энергичному росту онтостадийно более молодых корней. Если бы подвой был или оставался более стадийно старый, то привой с чрезмерным апикальным доминированием проявлял бы еще более высокую степень сенелитирования, например в виде полного отсутствия боковых ветвей и чрезмерностью роста главного = матрикального побега в виде плети с большим количеством шишек на прошлогодней его части. В итоге такая плеть быстро была бы сломана снегом или ветром.
Это еще раз подтверждает, что сенелит исходит, то есть его обуславливают часто не просто состояние апикальных почек, а и энергия роста сопряженных им противофазы – состояние корней.
Например, посмотрите фото с быстро постаревшими и отмирающими секвойями, где основной причиной их отмирания являются недостаточные почвенные условия для деревьев гигантов. Это привело к ухудшению работы их корневой системы, их недоразвитости, а затем ускоренному отмиранию рощи. Выраженная недостаточность апикальной активности корневых меристем является определяющим фактором для взаимоувязанных с ними верхушечными апексами. Корень и верхний апекс – это два противоположных полюса единой системы определяющих = обуславливающих состояние друг друга.

На фото: Изображение ели-мутанта с исключительным преобладанием плагиотропных побегов, форма «пендула» = плакучая и отсутствием ортотропных побегов.
В норме обычно плагиотропность растений сопряжена с наиболее высокой стадийной продвинутостью и продвижением к сенелиту. Но в данном случае этой связи не имеется, как и нет типичных признаков сенелита, когда ограничен рост и чрезмерно развита матура с признаками усыхания ветвей. Такой фенотип связан не с возрастным элевационным процессом = гормональными перестройками, а с банальной мутацией без продвижения по ступеням стадийности.

На фото: Старый дуб на вершине каньона на скальной почве с ограниченным ростом; побеги и листья по строению ближе к ювенильным особям с недозакладкой желудей. Регулировка фенотипа и онтостадийности управляется сугубо внешними суровыми факторами и гибелью апикальных почек.
Здесь пример приспособленческой индивидуальной реакции на самовосстановление ювенильности.
Изложенный ракурс позволяет говорить о возможности самоомоложения за счет преимущества отрастания из почек с более нижней стадией зрелости – адальтуса, но не матуры. Следовательно, если говорить о стволовых клетках, то стадийная зрелость здесь будет на несколько ступеней ниже, чем у развитых дифференцируемых тканей, но в тоже время выше, чем у тотипотентных и плюрипотентных. Таким образом они обычно проявляют промежуточную стадийность, но при этом несколько ниже, чем у генеративно детерминированных,
Самоклонирующиеся виды растений проявляют максимально высокую продолжительность жизни.
К таковым можно отнести, например один из видов елей, которая способна давать вегетативные отпрыски вплоть до 9550 лет. Особенностью её является обитание в холодных арктических условиях, когда верхняя часть дерева постоянно обмерзает и оно развило в себе способность давать вегетативные отпрыски, но из растущих плагиотропно = стелющихся нижних веток, которые зимой снег заново прижимает к земле, что даёт им способность перезимовывать под глубоким снегом в виде так называемых хвойных стланцев, например в тундре. При этом растение приспособилось давать отпрыски из брахиальных = пазушных или из спящих почек. Стадийно такие почки более ювенильны и не способны в этот же год закладывать генеративные побеги и шишки. Из-за плагиотропности, который поддерживается топофизисным свойством, рост таких побегов всегда замедлен и поэтому они более устойчивы к морозам в отличие от побегов из апикальных. Таким образом, сланцы всегда более стадийно молоды и морозоустойчивы. Это обеспечивает им высокую регенеративную способность и позволяет отключить истощающую растение фазу матуры = перезрелости и закладки чрезмерного количества шишек.
На фото видно, что дерево живёт в крайне тяжелых почвенных условиях, то есть практически это сплошной камень и мох. Ни о каких быстрых темпах роста здесь не может быть и речи, как это было на примере секвойи. Эта низкая вегетативная активность зафиксирована генетически и жизнеспособность не привязана к темпам роста, а к способности отрастать из брахиальных почек.
Тем не менее, дерево приспособило = переформатировало свою биологию, свой тип выживания, длительную ювенильность и адальтус к этим трудностям в росте и продолжает проявлять свою жизненную силу в самых трудных условиях арктики. Это пример реализации другого типа поддержания длительной молодости. Осуществляется это за счет других механизмов: не высокой вегетативной активностью, а за счет регенерации из стадийно более молодых почек – брахиальных, но из плагиотропных побегов. Постоянное самообновление дает ряд преимуществ: повышает морозостойкость, увеличивает потенциал длительности жизни до тысяч лет и не допускает до стадии матуры, а значит и до сенелита.

На фото: Ель, ствол которой имеет возраст несколько сот лет, но при этом она развила уникальную способность самоклонироваться, что позволило растянуть способность выживания её до 9500 лет.

На фото: столоновый побег способный к непрерывному восстановлению новых генеративных растений и вегетативных столонов, позволяющих растению непрерывность и неограниченность существования.
В междоузлиях растения заложены зачатки корней и будущих побегов. Жизнеспособность таких столоновых зарослей неисчерпаема. Растение за год может быть несколько раз скошено, но продолжать активно занимать ареал, сохраняя высочайшую жизнеспособность и молодость.

На фото: Феномен одного супердерева Тополя осиноподобного в виде клональной колонии, напоминающей рощу, на площади 43 га и весом 6000 тонн чистой массы. Растение распространяется вегетативным путем корневыми отпрысками. Предположительно возраст этой рощи около 80000 лет.

На фото: Слева обычные побеги криптомерии японской, а справа побег от ювенильной формы не способной закладывать репродуктивные побеги со стробилами.

На фото: Криптомерия японская ювенильная форма того же дерева. Генеративные мужские побеги недоразвиты и поэтому плохо образуют фертильные шишки.

На фото: Криптомерия японская внешний вид древа во взрослом состоянии.

На фото: Внешний вид обычного хвойного дерева ели.

На фото: Внешний вид того же вида хвойного дерева ели, но ювенильной формы, то есть проявление неотении.
В природе встречаются случаи, когда ювенильные особи хвойного растения дают отрастать побегам аналогичным как у нормальных растений. Это так называемое обратное перерождение, что говорит о том, что ювенильность у этих растений не связана с генетической мутацией. Чаще всего это локально размещенные почечные спурты, аномалии их развития, которые затем размножают вегетативно. Они могут представлять собой биологические химеры, когда в одной ткани или клетках сожительствует несколько типов разных клеток. Например, пестролистные плющи в разных условиях могут израстать из своей пёстролистности, а в других возвращать свое состояние. Это своеобразная вариабельность.
В данном случае здесь можно говорить о закрепленной определенным образом ювенильности = неотении у древесных растений. В норме ювенильная форма побега и расположение хвои имеется только у сеянцев, а в данном случае этот признак сохранён во взрослом состоянии.
Почему мною это так подробно рассматривается? Ситуация здесь аналогична тому, когда мы различными методами сможем навести организм человека в состояние неотении, но даже после этого необходимо понимать как это состояние можно поддерживать длительно, как не потерять достигнутого и какие механизмы этим управляют. Только на примере растений всю эту закономерность можно подсмотреть.
Очевидно, такой возврат из ювенильного состояния роста побегов во взрослые связано с особенностями расположения побегов в кроне и их разнокачественности. Очевидно, те участки верхней части кроны, где апексы побегов наиболее хорошо обеспечены определенным спектром гормонов, способны к запуску в них генетических программ обычного развития. Следовательно мутация связана с недостаточной чувствительность к гормональному фону в юном возрасте, и эта программа зрелости не запускается, но тем не менее на определенных запоздавших этапах развития такое переключение программ на новый этап перехода во взрослое состояние восстанавливается.

На фото: Внешний вид ювенильной формы хвойного дерева ели, на вершине которого выбились нормальные побеги. Возобладала нормальная меристема почек.
Объяснение механизма ювенильности на примере плюща.
Состояние ювенильности может быть бесконечным и принципиальных препятствий этому нет.
Удобной моделью для объяснения механизма ювенильности является плющ обыкновенный. Особенностью этого растения-лианы является то, что на одной особи одновременно могут находиться и онтостадийно зрелые ветви с плодами и ювенильные побеги. Зрелые с плодами ветви располагаются в верхней части растения, где больше света, а нижние с ювенильной формой расположены внизу в более затемнённых местах. Фаза ювенильности может быт растянута у растения чуть ли не до бесконечности, до тех пока растение не наберёт нужной биомассы, а верхние ветви на получат достаточной степени фотопериодическую индукцию.
На фото: Ювенильные и взрослые плодоносящие побеги взятые с одного растения.
Ювенильные побеги у плюща никогда не дают образование плодов, последние образуются только на морфологически зрелых укороченных ветвях. У ювенильных побегов листья вырезанные как у клёна и имеется большое количество корневых зачатков, побеги вытянуты, легко укореняются, а у зрелых – листья овальные.
Стадийная реювенилизация реальная возможность в растительном мире.
Пример с плющом доказывает возможность обратного развития по реостату стадийного онтогенеза.
Почему это важно осознать? Рассматривая пример размножения клоновыми черенками секвойи из кроны, мы отмечали, что они не способны отменить заданные им в кроне особенности развития и роста, проявляя топофизис = морфологические особенности строения побегов в кроне и циклофизис = способность к плодоношению.
Онтофизис = онтостадийность - принципы и механизмы их регулировок.
Термин предложен Гарбузовым Г. А. – подразумевает не простое сохранение у клонов неких особенностей роста и плодоношения, а проявление более общего процесса, то есть сохранения нового качества - степени онтостадийной зрелости. При этом расширено понимание этого явления не только для клонов, но и в пределах данной особи растения, когда новое поколение побегов в пределах одного и того же дерева все проявляют продвинутую физиологическую или морфологическую стадийную зрелость. Программы на ювенильность у них заблокированы. Онтофизис это пример продвижения по лестнице этапов онтогенеза, когда приобретенная зрелость закрепляется навсегда как в клонах, так и последующих побегах в пределах растения. В животном мире под этим будет пониматься сохранение у потомства линий клеток только тех возрастных стадийных качеств, которые достигли им предшествующие клетки.
Если привить взрослую форму плюща на ювенильную, то у взрослой формы можно наблюдать возврат к образованию ювенильных листьев, закладки корней, закрытию способности на онтофизис.
В свою очередь можно ожидать, что если привить ювенильную форму плюща на зрелые побеги, то это приведёт к переводу ювенильности на стадийную зрелость. Следовательно, каждая стадия развития или зрелости выделяет вещества, которые только ей присущи, которые сами в себе и поддерживают своё состояние.
Еще раз подчеркну, что здесь нужны обязательно изменения не только на верхнем уровне гормонов и т. п., но и на уровне генов. Для этого нужны разные программы. Настройка уровня гормонального фона должна осуществляться своей программой, а для экспрессии генов перехода на очередную онтостадию - другая программа. Перестройки должны закрепляться на всех этажах сразу. Первоначально верхний этаж со своим набором программ контроля гормонального фона и соответствующего ему роста создаёт условия необходимые для нижнего этажа, когда становится возможным переключение на новые блоки.
Когда на нижнем генном этаже осуществлена перестройка, то клоны или линии клеток от их источника, то есть за пределами маточного исходного растения или животной особи, всё равно продолжают сохранять статус и уровень онтостадийности за счет генопрограмм нового экспрессированного блока. Последние могут не отключаться у клонов. Закрыть эти программы можно только экспрессированием более ранних программ ювенильности.
Не устойчивый онтофизис. В то же время клоны, которые получены от особей с изменениями связанными только с верхними этажами гормональных регулировок, без закрепления на нижних, могут не поддерживать = сохранять онтофизис и вернуться на прежнюю онтостадию.
Вывод. Для чего важно понимать эту двухэтажность регулировок? Например, если мы хотим противодействовать как сенелиту, так и матуре и адальтусу с целью возврата в ювенил, которые являются полноценным проявлением двухэтажных преобразований перехода на каждую онтостадию, то все наши действия по коррекции верхнего гормонального этажа будут недостаточны и после их отмены приведут к возврату в эту стадию. Большинство существующих экспериментов по преодолению сенелита направлены только на одну сторону этого процесса и как итог вывод, что сенелит это уже навсегда.
Реювенилизация или обратный онтогенез возможны.
Эксперимент на плюще позволяет сделать очень важный биологический вывод: у ряда видов растений имеется возможность возврата от стадий сенелита и матуры в адальтус и ювенил = реювенилизации. Ранее такой возврат считался не возможным, поскольку онтогенез может двигаться якобы только в одну сторону.
Итак, на примере плюща мы видим реальную возможность стадийного омоложения. Принципиальных преград этому нет. В мире растений, в частности у плюща, легко осуществляется возможность избежать этапа старения. Какая программа онтогенеза заложена, такая и реализуется. Даже если у организма нет потенций и программ на непрерывное существование, то в это отсутствие соответствующей программы можно вмешиваться коррекциями из вне по переходу, возврату на ювенильность, а точнее пролонгирование неотении.
Факторы развития – химические и физические как движущая сила.
Двигающими силами перехода на новый этап развития могут быть как изменения гормонального фона связанного с непрерывным ростом, так и ряд факторного воздействия, например: длина светового дня, влажность, температура, полное отсутствие света = этиоляция и другие. То есть на верхнем этаже регулировок работают как гормоны, так и некие факторы регуляторы роста и развития.
Фотопериодические механизмы онтостадийных регулировок.
Удобной моделью для исследований по онтогенезу является Дурнишник. Особенностью растения является чёткая фотопериодическая реакция, то есть способность растения зацветать в ответ на определённую длину светового дня (фотопериод). Световой день весной и осенью короче, чем в середине лета. У растений есть специальные механизмы, обеспечивающие специфическую реакцию на фотопериод. У длиннодневных сигналом для образования цветков служит удлиненный летний день, короткодневным для этого необходим укороченный световой день. Дурнишник строго короткодневное растение. Он может оставаться в вегетативном = ювенильном состоянии как угодно долго и не цвести, если его выращивать в условиях длинного светового дня. Взрослые растения перейдут к цветению даже при одном укороченном световом дне. Но в юном возрасте, когда только развернулись семядоли, растения не зацветают и при 10 коротких днях. Тем не менее, уже первым двум листьям, следующим за семядолями, достаточно одного короткого дня, чтобы индуцировать образование зачатков цветков. Таким образом, просматривается хорошо, что индуцирующий фактор может проявить себя только в подготовленных для этого тканях. Такая подготовка, очевидно, связана с предварительным изменением гормонального фона, что возможно только в новых листьях.
За ростом этого фона следят датчики генопрограмм. Они контролируют уровень мощности действия гормонального фона. Когда этот уровень, фон достигает достаточных величин, то ихние программы, заблокированные программами ювенильности, проявляют готовность к экспрессиям. То есть экспрессия предварительно должна быть подготовлена. Для этого нужны верхние этажи регулировки и механизмы их реализации.
Это в свою очередь обеспечивает готовность к онтостадийному продвижению, а последнее в сочетании с индуцирующим фактором обеспечивает нужное переключение и экспрессию генов.
Двухэтажность регулировок стадийного перехода: это обязательно внешний количественный этаж, то есть общеорганизменный уровень становления гормонального профиля, что в последствии позволяет сработать нужному фактору для закрепления процесса и перехода на второй этап, то есть генетический = новый программный уровень.
Двуэтапность онтостадийного продвижения.
Такое сочетание изменений, согласование процессов как на верхнем уровне, так и на генетическом уровне, которое приводит к закреплению, фиксированию его на программном уровне, то есть экспрессии иного комплекса генов, и есть этап онтостадийного продвижения, то есть нового устойчивого физиологического состояния. Подчеркну, что изменения здесь происходят на обоих уровнях регулировки сразу.
Верхний уровень регулировки имеет свои ограничительные механизмы. Например, процесс роста имеет коридор свободы только в определенных границах. Это саморегулирующийся гомеостат, которым управляет своя система программ. Гомеостат этот определяет параметры коридора. Все они зафиксированы генетическими отдельными программами. Когда верхний уровень коридора свободы роста заполнился, то включаются генетические механизмы его ограничения. Бесконечным рост у высших животных не может быть. Он всегда корректируем. Это является началом второго этапа генетических перестроек на нижнем этаже.
Итак, первично включаются ограничительные механизмы, включающие переформатировку гормонального профиля. Это этап № 1. За этим этапом включается этап № 2 – генетических перестроек = подстроек.
У Дурнишника оба механизма регулировки цветения могут находиться на одном растении и даже в одно и то же время. Семядоли здесь можно рассматривать как органы с ювенильными свойствами, активно противодействующие цветению, а первую пару листьев - как стадийно продвинутые.
Семядоли Дурнишника не только слабо восприимчивы к фотопериодической индукции, но и способны ингибировать начавшийся переход к цветению. Очевидно, листья 1-й пары, с высокой восприимчивостью к фотопериодической индукции, являются источником гормона цветения.
Следовательно, дурнишнику так же как и плющу, ювенильность обеспечивает какое-то вещество, способное подавлять цветение. Однако, эта способность снижается с возрастом. Онтостадийность его блокирует, а точнее блокирует ювенильность, когда этапы роста полностью исключены.
Мортальный фактор.
Дурнишник, в отличие от плюща, растение с 1-летним циклом жизни, где четко прослеживается фактор ограниченности его жизни, то есть мортальный фактор, который жестко связан с наступлением в онтостадию адальтуса и сенелита одновременно. Правомочно ли здесь говорить о сенелите, когда растение после плодоношения включает абсцизовую кислоту, которая его и убивает? Сенелит не обязательная стадия развития. Мортальность не обязательное условие жизни.
Иммортальность онтогенеза.
В свою очередь растение плющ по сути иммортально = бессмертно, так как способно поддерживать не ограниченно стадию ювенильности.
Онтостадийность как фактор предрасположенности к возрастным болезням и механизмам мортальности.
Онтостадийные изменения приводят не только к изменению чувствительности клеток к различным патологическим факторам, но и к тому, что на разных онтостадиях могут проявляться различные заболевания. Этим можно объяснить, что у животных на стадии сенелита появляется свой круг заболеваний. Например, те же самые ткани в молодости не подвержены ряду заболеваний, тогда как в сенелите эти проблемы становятся закономерными. Например, мыши в молодости не страдают от недостатка аминокислоты метионин, тогда как для взрослых особей это ведёт к предрасположенности к провоспалительным процессам. Первый способ бороться с воспалением путём усиленной подачи метионинсодержащих продуктов, а второй тормозить онтостадийную зрелость и эта проблема сама собой решиться.
Поэтому бороться с болезнями старости у человека правильнее не путем противодействия каждой из них по отдельности, а путём отодвижения стадий матуры и сенелита.
Вывод: онтостадийные перестройки – несут в себе генетическую основу для болезней возраста.
Онтостадийные перестройки на первичном генном уровне важное условие возрастных болезней. Дело в том, что многие теоретики слишком много внимания уделяют второстепенным причинным факторам старения, например закисленность, повышенная воспаляемость, слабая клеточная энергетика, нарушение работы митохондрий, укорочение и не восстанавливаемость длины теломер, и другие, которые выдаются за первопричины проблемы. Но все они являются следствием перехода на очередной онтостадийный этап, закрепленный в онтофизисе, позволяющий триггерно проявиться этому вторично проявленному симптомокомплексу болезней сенелита. Этот симптомокомплекс сцеплен с онтостадийностью, что означает, что лечить эти болезни практически не возможно без одновременного снятия программ сенелита и адальтуса.
Гормональный профиль ювенильности и адальтуса у растений.
Точно также, содержание ауксинов в семядолях дурнишника было выше, чем в листьях. Обработка раствором ИУК растений дурнишника также подавляет образование зачатков цветков. Следовательно, у дурнишника, ауксин действует так же, как у плюща. Обнаружено, что обработка ауксинами еще не цветущих, но близких к этому растений, задерживает зацветание. Ауксины и есть тот внутренний фактор, не позволяющий растениям цвести.
Но этому противоречат данные, показавшие, что более поздняя обработка растений ауксинами, когда цветение уже началось, его, напротив, усиливает. Это означает, что физиологические процессы растений регулируются несколькими гормонами роста. Имеются разные типы роста, например: деление клеток и их растяжение, рост в длину или рост в ширину.
Роль цитокининововых гормонов.
В семядолях дурнишника уровень цитокининов так же как и ауксинов был выше, чем в листьях. Нанесение цитокининов на индуцированные к цветению растения подавляет цветение, так же как это делает ауксин.
Роль гормона торможения роста абсцизовая кислота.
Среди гормонов роста растений имеются не только регуляторы, активирующие разные стороны роста, но и задерживающие, подавляющие его; один из таких негативных гормонов роста ‒ абсцизовая кислота. Это гормон, регулирующий опадение листьев осенью. АБК накапливается также при созревании семян. Это предохраняет их от преждевременного прорастания. Содержание АБК увеличивается при засухе. Иногда она накапливается и в интенсивно растущих молодых листьях. Интересно, что при засухе молодые листья погибают последними, после того, как засыхают взрослые листья. АБК накапливается и при переходе к цветению. У некоторых растений обработка АБК может вызвать образование цветков. В семядолях дурнишника обнаружено лишь следы АБК
Напротив, в листьях её содержание было высоким. Однако самое высокое содержание АБК оказалось в молодых растущих листьях. Они, кстати, оказались и самыми чувствительными к индукции цветения коротким днём. Так что содержание АБК у дурнишника больше коррелирует не с задержкой роста, а с чувствительностью к фотопериодической индукции. Обработка семядолей АБК на фоне фотопериодической индукции позволила большему числу растений образовать зачатки соцветий. Все эти результаты показывают, что способность семядолей ингибировать переход к цветению опосредована высоким содержанием гормонов, усиливающих рост, и низким содержанием гормона, задерживающего рост. Таким образом, ювенильность растений находится под полигормональным контролем.
Гиббереллины – гормоны участники регулировок роста и вытяжки стебля.
Растения с длиннодневной фотопериодической реакцией обычно до перехода к цветению образуют короткие, сближенные междоузлия. Заложение зачатков цветков и соцветий сопровождается усилением роста междоузлий, вызываемое гиббереллинами. Низкий уровень гиббереллинов у длиннодневных растений может быть одной из причин ювенильности. Обработка гиббереллинами вызывает образование цветков. У растений с потребностью в яровизации ювенильное состояние также обусловлено недостаточным для зацветания уровнем гиббереллинов. Если гиббереллины дать извне, то растения будут цвести. Выраженные результаты об участии гиббереллинов в регуляции перехода от ювенильного ко взрослому состоянию были получены на хвойных деревьях. Обнаружено, что гиббереллины, которые впрыскивали с помощью шприца в древесину, способны вызвать раннее цветение у многих видов семейства Кипарисовых и Сосновых. Важно, что с прекращением обработки гиббереллинами, хвойные деревья возвращаются к ювенильному состоянию. Очевидно здесь наблюдается не устойчивый онтофизис. Похоже, это связано с неким неопределённым внутренним фактором поддержания ювенильного состояния, который возвращает деревья в ювенильное состояние после прекращения действия гиббереллинов. Простое наличие гиббереллинов недостаточно, чтобы переключить генопрограммы на устойчивый переход на онтофизис. Эта способность возвращать в ювенильное состояние используется в лесоводстве: перспективность хвойных деревьев обнаруживается поздно, когда они становятся взрослыми. Однако черенки со взрослых деревьев плохо укореняются и не годятся для размножения. Поэтому для укоренения их надо сначала омолодить, возвратить в ювенильное состояние. С этой целью верхушки взрослых деревьев последовательно перепрививают несколько раз на ювенильные проростки.
Обнаружено у позднеспелых сортов гороха, с удлиненным ювенильным периодом, специальные гены, обусловливающие задержку перехода к цветению и ответственные за продуцирование ингибиторов цветения. Поздние сорта характеризуются активным, доминантным состоянием генов ювенильности. Такие растения становятся фотопериодически чувствительными. В условиях благоприятного для цветения длинного дня уровень ингибиторов снижается и цветы закладываются после образования 15-17 листьев. В условиях короткого дня цветы образуются после 20 и более листьев. Напротив, у ранних, скороспелых сортов эти гены несут в себе мутации, делающие их рецессивными (не проявляющими своё действие) и, следовательно, неспособными продуцировать ингибиторы цветения. Такие растения зацветают после образования 5-6-ти листьев независимо от фотопериодических условий.
Какова природа ингибитора цветения?
Если ювенильное состояние, как, например, у плюща, сопровождается повышенной способностью образовывать корни, то вероятно тот внутренний фактор, который вызывает образование корней, может также обусловливать и неспособность ювенильных растений зацветать. За образование корней ответственна особая группа гормонов роста – «ауксины».
Следует отметить, что онтостадийность должна здесь регулироваться двумя противоположными рычагами. Везде все типы тканей имеют два противоположных начала. В дальнейшем мы этот же момент будем рассматривать на различных тканях с противоположно направленными механизмами регулировок у тканей животных.
Ингибиторные и активаторные регуляторы цветения и ювенильности – факторы транскрипции.
В регуляции ювенильности принимают участие не только негативные регуляторы цветения, но и позитивные, которые ускоряют зацветание. Таковым оказался белок ФТ1, способный передвигаться по сосудистым путям растений и даже через место прививки. Более того, он оказался универсальным индуктором цветения для всех растений. Однако активность гена ФТ1 оказалась очень низкой у растений с длительным ювенильным периодом, особенно у деревьев. Введение в них генетическими методами гена ФТ1 значительно сокращало их ювенильный период и приводило к раннему цветению и плодоношению. Напротив, мутации, дезактивирующие, повреждающие ген ФТ1, отдаляли зацветание, увеличивали длительность ювенильного периода. Таким образом, недостаток гормона цветения ФТ1 является другой причиной ювенильности. Таким же универсальным оказался и ген (белок) с противоположным действием - ТФЛ1, ингибирующий цветение у Арабидопсиса. Он был выделен из многих растений, в частности, однолетних с относительно коротким ювенильным периодом, как уже упоминавшийся горох, арабидопсис, и других, а также из многолетних древесных растений с очень длительным ювенильным периодом: яблони, цитрусовые деревья и др. Мутации, делающие этот белок не активным, значительно сокращали длительность ювенильного периода. Таким образом, длительность ювенильного периода у растений обусловливается соотношением двух активных и противоположных групп факторов: положительного по отношению к цветению и отрицательного. Следует учесть, что гены ФТ1 и ТФЛ1 и их белки обусловливают вилку – вегетативный или репродуктивный, половой путь развития. Оба эти белка, сопровождаются группой других регуляторов, играющих важную роль в реализации выбранной программы развития. Среди них находятся и гормоны роста.
Следует также отметить, что сами белки генов ФТ1 и ТФЛ1 относятся к «факторам транскрипции». Большинство генов кодируют белки, выполняющие разные биохимические функции. Это ферменты, мембранные белки и др. Функция же «факторов транскрипции» совершенно другая – они включают, активируют считывание генов. В процессе считывания гена (transcription) принимают участие многие белки и сам ген довольно сложно устроен. Мы выделим лишь то, что связано с действием факторов транскрипции. Ген, грубо говоря, можно разделить на две основные части. Во-первых, это сама программа для синтеза определенного белка, записанная в ДНК. Она называется «открытая рамка считывания», ОРС. ОРС считывается ферментом РНК-полимеразой и затем воспроизводится в виде мессенджер 9 РНК (mRNA). Она после структурных изменений (splicing) переписывается (процесс называется транскрипция) в конечный белок. Другая часть гена – регуляторная. Это «промотер». Пространственно он предшествует ОРС. Именно его должны обнаружить особые белки - факторы транскрипции. Они должны сесть на промотер, обозначить место для посадки РНК-полимеразы и запустить ее работу. Считается, что гены ФТ1 и ТФЛ1, регулирующие ювенильность, не только факторы транскрипции, но и гены-интеграторы. На них сходятся регуляторные цепочки других процессов, участвующих в регуляции возраста, в частности ювенильности.
Роль апикальной меристемы в онтофизисе.
У растений инициация и рост органов, листьев и цветков, происходит в специализированных местах – в точках роста = апикальных меристемах, апексах. На гистологическом срезе выявляются специализированные клетки апекса. Так на срезах апексов можно выделить участки стволовых клеток, которые дают начало клеткам с разной специализацией. В них активированы гены, ответственные за эту специализацию. У ювенильных растений апекс маленький и ему надо дорасти до определенного размера, чтобы среди его клеток произошло разделение по функциям.
Включение стадии матуры = сверхзрелости может быть стартерным механизмом для сенелита.

На фото: Сельдерей в 1-летнем возрасте не проявляет образование цветочных почек и находится на ювенильной фазе.

На фото: Видно что переход растения сельдерей на второй год в стадию плодоношения вызывает у него пожелтение и быстрое старение листьев, а затем отмирание.
Итак, у многих видов 1- и 2-летних растений наступление стадии плодоношения = адальтуса, равнозначна губительной стадии матуры = сверхзрелости у многолетников, жестко сцеплена с генопрограммами включения сенелита и самоуничтожения растений. Одна программа обуславливает наступление другой.
Особенности ювенила у многолетних, но монокарпических растений.
Отсутствие предшествующей программы часто может отменить фиксированный сенелит. Таковой является например агава. Она даёт единственный генеративный побег на 8-15 году жизни, то есть цветет один раз в жизни. Тем не менее, если не давать растению цвести путём обрезки цветоноса, может жить до 100 лет и погибнуть не из-за стадийной старости, а из-за механистических внешних факторов израстания за пределы возможностей для дальнейшего роста, то есть выход за оптимум для её жизни = коридора гормезиса.
Следовательно, здесь проявлен жестко фиксированный онтогенез.
ОСОБЕННОСТИ РЕГУЛИРОВКИ ДЛИТЕЛЬНОСТИ ЖИЗНИ В ЖИВОТНОМ МИРЕ.
Низшие животные могут не стареть и не умирать.

На фото: Бессмертная медуза Turritopsis.
Морские медузы могут внезапно омолодиться. Процессы в организме этих созданий могут обратиться вспять. И медуза вновь возвращается в состояние подростка.
Эти существа показывают, что старение и смерть — биологически не заложенная неизбежность.
Морские красные ежи - это пример, когда организм никогда не стареет. Они живут, растут, размножаются, но не меняются, оставаясь такими же молодыми, как были. Их вечная юность может продолжаться и сто, и двести лет. Возможно, что и больше.
Гидра способна в нужный момент обновить свои клетки. И снова омолодить свой организм. А ведь обновление стволовых клеток — не только путь к молодости, но и способ вылечить многие болезни. Гидра технически бессмертна: они погибают только от врагов или неблагоприятных условий, но не от старости. Она же обладает высшей способностью регенерировать. Даже из нескольких клеток может вновь вырасти целая гидра. Если все хорошо – особи размножаются половым путем, если что-то пошло не так – самостоятельно выращивают своих клонов.
Итак, гидра может жить без ограничений. Здесь и кроется ответ, что и в животном мире первичное старение на клеточном уровне не обязательно.
У низших животных первичное старение, связанное с клетками необязательно и не неминуемо, а закладывается филогенетическими необходимостями.
Таким образом, мною приведено все больше и больше данных, что старение и в животном мире не обязательно, не является неотъемлемым свойством живого, а эффект Хейфлика ни о чём не говорит, а всего лишь одна из программ, которая больше связана с блокировкой предшествующих программ, то есть аналог клонового циклофизиса, которое описано мною выше. Через эти механизмы эволюция управляет темпами смены поколений: когда ей это целесообразно программы по блокировке к реювенилизации отсутствуют.
Губка - рекордсмен по долгожительству. Эти животные, похожие на растения, обитают по всему миру в соленой и пресной воде. В 1986 году была найдена особь возрастом в 10 000 лет.

На фото: Стеклянная губка Euplectella aspergillum.
Стеклянные губки по расчетам ученых живут по 15000 лет. И нет достоверных сведений о том, умирают ли они вообще своей смертью. Стеклянные губки могут замедлять и ускорять процессы старения без видимых причин. Их внешний скелет, напоминающий стекло, надежно защищает их от врагов. Феноменом старения они не обладают.
Раковины европейской жемчужницы могут жить по 400 и более лет.
Омары в принципе бессмертны. Они способны вырабатывать фермент, замедляющий старение. Это бесконечно растущие существа. Самый большой омар, выловленный людьми, весил 20 кг и ориентировочно имел возраст около 200 лет.
Позвоночные животные могут обладать только долгожительством.
Галапагосские слоновые черепахи живут около 170. Описан случай жизни 250 лет.
Гренландские полярные акулы в среднем живут по 200 лет, но известны и 400-500-летние особи.
Гренландские киты доживают до 200 лет.
Преодоление онтофизиса в мире животных.
В экспериментах по парабиозу с животными при сшивании сосудистой системы молодых крыс к старым приводило к таким же результатам: старая особь проявляла все признаки омоложения, в том числе преодолевала онтостадийные преграды – онтофизис? Под снятием онтофизиса здесь понимается возврат стареющих линий стволовых клеток в изначальные. В свою очередь имеется и эффект обратного действия, когда старая особь вызывала ускорение старения у пришитых молодых. Значит имеются как вещества связанные с молодостью, так и вещества определяющие старение. Конечно, это может быть не просто наличие или отсутствие тех или иных веществ, а в большей степени связано с уровнем, порогом действующих веществ или их сочетанием, что мы обозначаем как гормональный профиль + профиль действующих факторов.
В целом можно утверждать, что та особь, гормональный профиль которой будет сильнее, в конечном итоге и окажет своё преобладающее действие. Если гормональный фон молодой особи будет слабее много более взрослой особи, то эффекта омоложения у старой почти не будет, но и более молодая быстрее состарится. Причем, эффект здесь сработает на уровне преодоления онтостадийного продвижения в ту или иную сторону.
В тоже время следует отметить, что в мире животных определяющими являются не гормональный фон, а некие факторы крови молодой особи, которые активируют генетические механизмы в стволовых клетках старого животного. Таковыми очевидно являются компоненты, ответственные за эти явления, а именно фактор дифференцировки роста GDF11.
Этот фактор, точнее сигнальная молекула необходима для нормального роста и формирования тканей, но с возрастом её продукция неуклонно снижается.
Переливание молодой донорской крови или инъекция фактора молодости, изъятого из крови – перспективное направление в омоложении.
Итак, сенелит обратим, а продление молодости реально. Для этого не обязательно «пришивать» к себе юного донора. Считают, что тот же результат можно получить путём инъекции GDF11 или переливанием молодой крови. Казалось бы фактор найден. Однако, попытки других исследователей по воспроизведению этих результатов не удались. Зато прижилась идея трансфузии и «кровяного эликсира».
В другом исследовании проводили пожилым крысам переливание плазмы (кровь, очищенную от белых и красных клеток), взятую у их 3-месячных сородичей. Это вызывало аналогичные результаты, что и при сшивании.
Гетерохронность старения тканей в организме.
В целях омоложения всего организма задача осложняется тем, что похоже каждый тип ткани идёт самостоятельно по этому пути и вносит свою лепту в этот единый процесс. Этим отличаются организмы животных от растений, у которых нет автономного уровня регулировок специфичной для каждого типа ткани. Организм животного это система из самостоятельных систем и тканей. Всё стареет в нём гетерохронно. Здесь старение гетерогенный = разнорусловый процесс, начинающийся в разных тканях заново, самостоятельно. В связи с возможной разной скоростью старения в разных тканях у разных индивидов происходит полиморфное проявление. Старение начинается на уровне каждого типа ткани самостоятельно и имеет свои индивидуальные внутритканные механизмы. Это как и при онкологии, когда каждый тип онкоткани несёт свой специфичный тип канцерогенеза и свои специфические механизмы.
Все типы тканей имеют разный уровень пролиферации клеток, например у клеток крови, слизистых оболочек он высокий, а есть ткани с нулевым уровнем митозов – нейроны, кардиомиоциты… Они прекращают деления еще в эмбриональной фазе. Нет митозов – нет срабатывания теломерного счетчика – нет укорочения теломер – нет ограничительного эффекта, а, например, кардиомиоциты от механизма теломер не должны стареть. Стареть должны быстрее клетки эпителия сосудов сердца, где идёт их интенсивная замена. Именно в них и описано проявление эпителиальной дисфункции сосудов с последующей гибелью части мелких сосудов и атеросклерозом крупных магистралей. Ключевым словом здесь является дисфункция = эпителиальная дисфункция.
В каждом типе ткани, системе имеются свои «инициирующие островки», которые и определяют свои механизмы клеточной дисфункции и свой путь к изнашиванию или старению.
Действительно любая ткань состоит из двух начал: функционирующей = работающей и противоположной ей, которая отвечает за регулировку первой, её трофику и индивидуальность клеточного восстановления. Похоже здесь применён механизм реципрокной регулировки: сколько клеток функциональной ткани ушло или износилось, столько должно и восстановиться с помощью управляющего начала.
Эти «инициирующие островки» не зависят от общего гормонального профиля и связанного с ним онтостадийного уровня старения. Существуют эти «островки» по своим внутренним законам и правилам, с помощью своей сигнальной системы регулировки и своих факторов роста, что и отображается на их особенностях принципов старения. Но можно ли предполагать что, это и есть первичный межклеточный уровень старения? Какова роль в этой схеме эффекта Хейфлика? Проще всего бы было предположить, что он и является определяющим ограничительным фактором существования данной клеточной системы. Но ниже будет приведена концепция Гарбузова Г. А., что ограничительный механизм Хейфлика, скорее всего, является механизмом противодействия нерегулируемым митозам. Но в условиях организма каждая ткань имеет свою тканеспецифическую генную систему контроля над митозами. Любые митозы здесь увязаны = сцеплены с механизмами провоспаления, которые и уравновешивают их. Это своеобразная триггерная система регулировок, когда любая инициация митозов возбуждает и открывает другие параллельные программы и при этом находится под строгим контролем ряда других программ. Таковыми в частности являются механизмы провоспаления, которые тормозят = ограничивают первые. Открытых = свободных = независимых систем, которые поддавались бы контролю с верхних этажей регулировок митозов, здесь нет.
В клеточных культурах ткани этот механизм провоспаления, очевидно, запускается благодаря тому, что нет противоположных начал: функционирующей и регулирующей стороны, система «открыта» из-за отсутствия реципрокных, противоположно направленных механизмов регулировки. Эта неуравновешенная система = неуправляемая, что в данной ткани всегда строго пресекается. В такой системе ускоряется процесс пролиферации, так как в условиях отсутствия тормозов система расценивает данную ситуацию как необходимость к регенерации. Это в свою очередь в нерегулируемой = неуравновешенной системе ведёт к чрезмерной скорости и количеству митозов. В условиях закрытой тканной системы это недопустимо, на пути этому природой предусмотрены буферные механизмы блокировки. Таковым ограничителем, работающим на «отсечение» данной линии пролиферантов, и является механизм Хейфлика. Любая тканеспецифическая линия клеток с таким нерегулируемым ростом должна пресекаться. Только линии клеток не достигшие уровня тканеспецифичности, то есть имеющие тотипотентные универсальные задатки, не пресекаются.
Такого эффекта Хейфлика нет в мире растений, он там не нужен, что даёт им возможность бесконечно клонироваться.
В животном мире другие принципы регулировок, которые опущены на межклеточный уровень. Механизмы открытого роста и самоклонирования здесь недопустимы. Любой рост должен быть скомпенсирован. Получается, что каждая ткань это в первую очередь система сама в себе. Тогда что в ней в пределах всего организма может запускать механизмы провоспаления, а значит и неограниченного или ограниченного роста и вслед за этим старения? Теоретически такая замкнутая реципрокная самовосстанавливающаяся система может существовать сама по себе неограниченно. А это есть залог того, что органы и ткани могли бы существовать в условиях организма неограниченно и не ограничивать своими правилами жизнь организма.
Тогда ограничения её возможностей должны исходить из уровней более высокого порядка. Напомню, что эксперименты по омолаживанию животных с помощью сшивания старых и молодых особей показывают возможность омоложения или старения на уровне этих тканей. Такая замкнутая автономная реципрокная система аналогична по своим возможностям с самоклонирующимися или непрерывно растущими открытыми системами у растений способных к неограниченному существованию. Следовательно, в обоих случаях систем открытых и закрытых старение не является окончательным = не обязательным процессом, то есть тот, который базируется на эффекте Хейфлика. В тоже время у растений показана возможность неограниченного существования, тогда как в мире животных имеются какие-то механизмы ограничений…
Теперь давайте посмотрим на этот же процесс старения, но с других позиций. Итак, исходим из посыла, что старение клеток контролируется изменениями длины теломер. Ключевой вопрос состоит не в том, насколько длинными были ваши теломеры при рождении, а в том, насколько короче они стали. Экспрессия генов меняется именно из-за укорочения теломер. А что если предположить, что их укорочение связано не только с механизмами деления, а и с механизмами провоспаления? Процессы количества митозов и внутриклеточных провоспалений приравниваются к друг другу. Тогда не ограниченное деление клеток в культуре ткани можно приравнять к провоспалительному процессу запускающему механизм Хейфлика. Допустим, в условиях организма провоспалительный процесс запускают как болезни, так и механизмы онтофизиса. Напомню случай описанный мною выше с мальчиком, который перестал стареть в возрасте 13 лет, и при этом у него проявлены все признаки неотении = ювенильности в возрасте даже 32 лет. Очевидно, остановка верхних программ, связанных с элевационными механизмами гормональных перестроек, обусловило остановку процессов провоспалений на межклеточном уровне и значит обнулило процессы тканевого старения.
Известно, что остановка старения у этого мальчика связана с нарушениями в работе гипофиза, который в свою очередь связан с гипоталамусом, а последний – с эпифизом. Можно предполагать, что именно эпифиз является дирижёром онтогенеза, который осуществляет свою работу через уровень мелатонинового нейромедиатора.
Но с этим не согласуются данные, когда пересадка молодой печени приводила у старого пациента к его омоложению. Этот случай указывает, что первичные механизмы общего старения находятся на уровне тканей.
Учитывая такую двойственность = неоднозначность выводов, мною предложено выдвинуть концепцию единства = параллельности программ движения процессов старения = развития верхнего общеорганизменного и нижнего тканевого, когда одно запускает другое? Становление определённых размеров и уровня работоспособности органов и тканей отключает ювенальный режим = статус, настройку верхнего дирижёра развития эпифиза. Достижение эпифизом нового элевационного уровня регулировки в свою очередь включает на этажах тканей программы по сохранению = фиксированию на геномном уровне в них достигнутого уровня онтофизиса = тканевую зрелость, продвинутость становления.
Очевидно, было бы более правильно обозначить два вида самостоятельных процессов, которые отображают уровни исходящих механизмов старения в организме:
- сенелит - верхний = гормональный = элевационный;
- сенесцент – нижний, межтканевой и клеточный.
То есть, старение идёт в двух направлениях одновременно с разных этажей становления организма. Одно запускает другое.
Иногда запускаются механизмы преждевременного старения:
- Senescentia praecox – связанный с нарушением гормоно-, нейро-, гуморальных осей регулировок.
- Прогерия - мутантный тип, связанный с нарушениями на генетическом уровне.
Сенелит – верхний уровень старения = гормональный = элевационный.
Элевационная гипотеза старения.
Дильман В. М. предложил эту гипотезу, в которой наиболее глубоко проработана сторона гормональных перестроек общего процесса старения. Также показаны механизмы неизбежности формирования возрастной патологии у высших организмов при переходах на завершающие этапы развития. Основную механизм перестроек он видел в возрастном повышении порога чувствительности гипоталамуса к гомеостатическим сигналам.
Особенность высших животных - наличие единой централизованной регуляторной надсистемы, которая определяет границы возникновения и развития в организме в процессе его онтогенеза различных гомеостатических систем. Это дирижёр всех процессов.
По сути это аналог апексов у растений, в которых тоже координируется работа всех гормонов. Но апексы в основном находятся в верхушечных побегах и через них определяется степень стадийного продвижения или зрелости побегов. Через апексы идёт реализация, воплощение необходимости.
В тоже время не апексы являются инициатором, первичным индуктором зрелости, а вся динамика сигнального фона обратных связей, то есть становления процессов на управляемых структурах. Аналогичные процессы идут и в гипоталамусе. Периферия задает условия центру.
Кроме регуляторных сигналов от желез внутренней секреции, гипоталамус корректируется и сигналами исходящими из нервной системы. В ходе этих процессов происходит возрастное включение и выключение функции репродуктивной системы в женском организме. Кроме того, одновременно происходят возрастные изменения в гипоталамо-гипофизарно-надпочечниковой системе, обеспечивающей тонический уровень глюкокортикоидных гормонов в крови, их циркадный ритм и повышение секреции при стрессе. В итоге, развивается состояние, которое можно описать как «гиперадаптоз».
Если перефразировать, то это более правильно обозначить как гипермодуляция = перекос одних гормонов над другими = перезрелость = матура = выход за пределы оптимального состояния и работы системы. Но развиваются эти процессы на платформе прежних онтофизисных = онтостадийных программ, что в итоге требует и перестроек на этом уровне. Здесь происходит не непрерывные количественные изменения = рост, а непрерывные качественные изменения. В пределах замкнутой модульной системы такие непрерывные качественные изменения рано или поздно выведут её из оптимального состояния, а значит и к энтропии, саморазрушению.
Действительно, стадия матуры = перезрелости с её климаксами не лучшее состояние для организма. Согласно этой концепции, вторичными являются перестройки, подстройки метаболического характера на клеточном уровне, которые якобы подстраиваются под верхний фон. В тоже время эта концепция не учитывает, что организм животного это интеграция из автономных = саморегулируемых систем и тканей. Всё стареет в нём гетерохронно, то есть это гетерогенный = разнорусловый процесс, начинающийся в разных тканях заново, протекающий по разному. В связи с возможной разной скоростью старения в разных тканях у разных индивидов происходит полиморфное проявление.
Таким образом, в разных тканях происходят одновременно перестройки как на нижних этажах регулировки, так и на уровне взаимодействия тканей. В тоже время это одновременно и двусторонний процесс пересекающихся потоков, где нижние уровни обуславливают работу верхних, а верхние влияют на нижние. Своеобразное взаимопроникновение процессов. Это обуславливает нарастание с возрастом содержания жира в теле, снижение чувствительности тканей к инсулину (предиабет) и развитие эпителиальной дисфункции – основы атеросклероза.
Элевационная теория устанавливает роль возрастных изменений, закономерно возникающих в трех основных "супергомеостатах": репродуктивном, адаптационном и метаболическом. В их пределах формируются ключевые по значимости для продолжительности жизни индивидуума способности: метаболическая иммунодепрессия, предрасположенность к возникновению злокачественных новообразований.
Концепция считает, что главные болезни, связанные со старением, не запрограммированы, а являются побочным продуктом реализации генетической программы развития, и поэтому старение возникает с закономерностью, свойственной генетической программе. Такая концепция упускает из виду, что в каждом типе ткани, системе имеются свои «инициирующие островки», которые и определяют свои механизмы клеточной дисфункции и свой путь к изнашиванию или старению. Первичные начала или ключи к старению в каждом типе ткани связаны с заблокированием на этапе эмбриональности и зиготы потенций к поной регенерации. Возможности клеточного уровня к полному восстановлению ограничены. Это означает, что односторонняя трактовка с помощью изменений гормонального профиля как главного = централизованного фактора старения ошибочно.
Да, любая ткань = система состоит из двух начал: функционирующей = работающей и обслуживающей её, которая отвечает за регулировку первой, её трофику и индивидуальность клеточного восстановления. Похоже здесь применён механизм реципрокной регулировки: сколько клеток функциональной ткани ушло или износилось, столько должно и восстановиться с помощью управляющего начала. Но этот принцип нарушается из-за предписанных пределов в клетках.
Эти «инициирующие островки» мало зависят от общего гормонального профиля и связанного с ним онтостадийного уровня старения. Тем самым, они вносят свой вклад с нижних уровней в общее русло сенелитных перестроек. В итоге это двусторонний процесс. Существуют эти «островки» по своим внутренним законам и правилам, с помощью своей сигнальной системы регулировки и своих факторов роста, что и отображается на их особенностях принципов старения.
Элевационная концепция связывает старение со старением гипоталамуса. Но гипоталамус это аналог апексов у растений, которым в принципе не обязательно присуще старение. В отличие от растений, у которых фаза репродуктивных процессов в старении может не угасать, а нарастать, у высших животных существует механизм отключающий её при старении.
Согласно элевационной концепции в гипоталамусе имеются чувствительные центры, которые реагируют на действие половых гормонов яичника по механизму отрицательной обратной связи. Иными словами периферия тормозит центр. С возрастом эта функция стимуляции гипоталамусом яичников ослабевает и ядро отдающее сигналы отключается. Клинически это подтверждалось тем, что у крыс среднего возраста снижалась чувствительность к эстрадиолу гипоталамических структур, ответственных за овуляторный выброс гонадотропин-высвобождающего гормона, что лежит в основе выключения репродуктивной функции = менопаузы. Это и есть наступление климакса = матуры. Для филогенеза не целесообразно, чтобы сенелитирующие ослабленные и больные особи продолжали репродуктировать потомство.
С позиций Г. А. Гарбузова это следует рассматривать как проявление закреплённого онтофизиса = продвижение в очередной онтостадийный этап развития = матура. Даже если репродуктивная функция здесь и закрывается, что казалось бы подобно ювенилу, тем не менее, это всё осуществляется на совершенно ином гормональном профиле, не благоприятном для особи.
Также как и у апексов растений, в гипоталамусе происходит смена стадийных генетических программ, перестройка. Это может проявляться в повышении порога чувствительности, что осуществляется путём уменьшения количества рецепторов клеточных мембран и снижения уровня нейромедиаторов гипоталамо-гипофизарного комплекса.
Гипоталамус - центральная мозговая структура, главный интегратор информации, поступающей из тела, и вместе с тем тот коллектор, куда вливается информация из окружающей среды.
По сути гипоталамус это централизованный штаб, пульт управления = интегратор = оптимизатор = коммутатор = триггер = координатор множества функций. Через гипофиз гипоталамус регулирует рост тела (гормон роста), деятельность щитовидной железы (тиреотропный гормон гипофиза), надпочечников (кортикотропин), функцию молочной железы (лактогенный гормон, или гормон, стимулирующий секрецию молока), репродуктивную функцию.
Ошибки элевационной концепции онтогенеза.
По сути, нет никаких элевационных процессов в гипоталамусе, а есть онтофизиные = онтостадийные перестройки всего организма, включая и первичные этажи с переходом на новые программы.
Стадийное развитие это единый процесс перестроек как на верхних этажах, так и на нижних генетических с переходом и фиксацией новых программ.
Итак, гипоталамус всего лишь опосредствующий механизм в роли рельсов для единого процесса перестроек на разных уровнях, на которых осуществляется регулировка. Этот интегратор не является системой самой в себе, первичным инициатором, индуктором, задающим развитие. Первичные начала стадийности задает не он, а всего лишь опосредует, осуществляет их реализацию. Истинные начала динамических перестроек находятся в регулируемых им тканях и органах мишенях. Отсюда идут сигналы обратной связи на необходимость перестроек в верхних этажах в т. ч. и гипоталамусе.
В концепции совершенно не учитываются механизмы онтофизиса = онтостадийности. Причиной матуры и сенелита является не гипоталамус. Он только посредник. Гипоталамус сопряжен с прилегающими к нему отделами мозга - ретикулярной формацией, где находится центр сна, а также центр, контролирующий эмоции и эпифиз, контролирующий циркадные ритмы. Эпифиз это дирижёр, а гипоталамус – партитура, оркестр, который разыгрывает на нотах центров аппетита, теплопродукции, теплорегуляции, роста, сна и бодрствования, эмоционального подъема, психической депрессии, размножения и многие другие. Его функции можно разделить на две группы:
Во-первых, гипоталамус приспосабливает деятельность организма к условиям внешней среды. Во-вторых, гипоталамус - это высший орган постоянства внутренней среды или гомеостаза. Вместе с регулируемыми органами гипоталамус работает, как своеобразная замкнутая система, обеспечивая постоянство внутренней среды благодаря механизму отрицательной обратной связи.
Стабильность внутренней среды, гомеостаз - обязательное условие свободной жизни организма. Однако одновременно в нем возникает непримиримое противоречие между потребностями роста, развития, размножения и необходимостью стабильности. Для осуществления всех этих потребностей необходимо нарушение стабильности, или увеличение мощности регулирующих систем.
"У высших организмов необходимо совместить одновременно покой и движение- покой внутренней среды, и движение, дающее развитие. Иначе говоря, системы, обеспечивающие стабильность должны сами развиваться, увеличивая свою мощность; только в этом случае может быть обеспечено сохранение регуляции в движущейся системе".
Согласно автору элевационной теории закон отклонения гомеостаза распространяется не на все регулируемые функции живого организма, а лишь на три из них. Эти три функции контролируют три основных свойства живого организма: способность живой системы к размножению, приспособлению (адаптации) и регулированию потока энергии (обмену веществ). Когда завершается выполнение программы развития организма, закон отклонения гомеостаза не прекращает своего существования, а, напротив, продолжает выполняться с той же последовательностью, что и раньше. Поэтому, если отклонение гомеостаза вначале служит развитию и росту, то затем оно превращается лишь в силу, нарушающую закон постоянства внутренней среды; после завершения роста развитие как бы продолжается и в результате постепенно начинают формироваться черты, свойственные нормальному старению и болезням старения. Действительно, в процессе старения нарушается гомеостаз: увеличивается в уровень сахара, холестерина и т.д. Следовательно, само нормальное старение есть болезнь, вернее сумма болезней, вызванных нарушением закона постоянства внутренней среды организма.
Увеличение мощности главных гомеостатов в процессе старения означает, что старение и связанные с ним болезни формируются не за счет снижения, угасания, а, напротив, усиления, перенапряжения деятельности систем, регулирующих энергетические процессы, адаптацию и размножение. Жизнь в процессе старения как бы идет вразнос, как рано или поздно нечто подобное произойдет с термостатом, если его регулятор из-за снижения чувствительности будет все менее и менее сдерживать повышение температуры в системе. Например, известно что сердце с годами слабеет. Но этому результату (ослаблению) предшествует увеличение его размеров, то есть усиление его мощности. Человек, старея, как бы движется по лестнице, ведущей вверх (поднимается по элеватору).
Возрастные изменения в деятельности трех основных гомеостатических систем - адаптационной, репродуктивной и энергетической - приводят к возникновению трех нормальных болезней старения - гиперадаптоза, климакса и ожирения, то есть болезней, которые с той или иной скоростью развиваются всегда в результате закономерного отклонения гомеостаза, связанного с осуществлением программы развития организма. Термин гиперадаптоз означает возрастные нарушения в адаптационной системе. Доказательством верности предложенной теории считал ученый то, что десять главных болезней из многих сотен возможных служат причиной 85% смертей. Ими являются: ожирение, сахарный диабет тучных, атеросклероз, гипертоническая болезнь, метаболическая (обменная) иммунодепрессия, аутоиммунные болезни, психическая депрессия и рак. Эти болезни, а также климакс и гиперадаптоз обозначены, как нормальные болезни старения.
Таким образом, на основе элевационной гипотезы старения появляется возможность искать взаимосвязь между главными и дополнительными факторами развития, созревания и старения организма, а также понять этиологию и патогенез болезней старения. Именно наличие перехода программы развития организма в механизм болезней старения даёт надежды в поиске путей и средств противодействия этим болезням. Никто не умирает от старости: люди и в старости умирают от болезней - в принципе излечимых, так как они вызываются нарушением регуляции. Естественная смерть у высших организмов - смерть регулярная!
Сенесцент – нижний уровень старения, межтканевой и клеточный.
Организм как саморазворачивающийся биомодуль.
Организм это саморазворачивающаяся многоуровневая система движущаяся по программам стадийного онтогенеза. По сути, это модуль-трансформер, который разворачивает свои возможности регулировок одновременно как с верхнего уровня, так и с нижнего. Каждый стадийный этап гормонального и факториального профиля является одновременно и шажком к трансформациям на геномном уровне = экспрессиям одних групп программ и репрессии других = скрытому «метаморфозу» = реорганизации рецепторного поля = структуры сенсорного дисплея на мембранах клеток + новый статус гормоно-, нейромедиаторного и гуморального профиля.
Любые общеорганизменные изменения отражаются и на межклеточном уровне, то есть обязательно фиксируются и на нижнем уроне регулировок, где тоже происходит переключение стадийных программ. Именно эта возрастная стадийность, трансформация сцеплена с теломерами. Чем дальше мы отодвигаемся от тотипотентности = эмбриональности, тем жестче и строже становится контроль над митозами. На уровне клеток это реализуется в виде фиксирования, геномного переключения перехода на новый режим, уровень регулировок закрепляющих приспособление клеток к данному онтофизисному состоянию. Это уже другие качественно и стадийно клетки. Программы клеток подстроились под внешнюю реальность и необходимость соответствовать им. Те же клетки, но работают на новом режиме.
В основе изменений возрастных перестроек на уровне клеток это фиксирование, достижение этого стадийного состояния. Это в свою очередь расценивается на уровне программ как ограничение = связывание их потенциальных возможностей.
Очередной виток уровня, усиления провоспалительных механизмов отражается и на уровне хромосом, что собственно говоря и проявляется в виде эффекта Хейфлика…
Единство провоспалительных начал хронической патологии и сенелита.
Известно, что любая длительная патология приводит к укорочению теломер. Как болезни могут повлиять на теломеры? Очевидно, через запуск провоспалительных процессов, а это сказывается на теломерах. Причем, процесс укорочения теломер происходит именно в обслуживающих клетках, но не в функциональных. Например, в глии, но не в нейронах, эпителиальных клетках сосудов сердца, но не в мышцах сердца. Происходит ли процесс провоспаления в самих функциональных клетках? Похоже, что нет. Но, тем не менее, здесь может происходить мощный дегенеративный процесс, который, очевидно, является вторичным после нарушений в регулирующем клеточном островковом слое из стволовых клеток. Именно они первично провоспаляются и стареют, а за собой тянут изменения в функциональных слоях.
Получается, что болезни и старение движутся на одних и тех же генетических рельсах.
Непосредственная и косвенная возрастная патология – в чём различия?
Учитывая, что нами предложено выделять три уровня старения в многоэтажном здании иерархии систем организма, целесообразно выделять непосредственную возрастную патологию, исходящую из нижних уровней, то есть на уровне старения клеток и косвенную, исходящую из верхних уровней, или когда стареющие клетки вызывают патологию в других тканях или клетках другого типа. Например, болезнь Альцгеймера и сердечно-сосудистые заболевания – проявления косвенной патологии, где нейроны и кардиомиоциты исполняют роль жертв патологических процессов в других компонентах системы.
Где находится общий знаменатель всех возрастных болезней?
Выше мы уже говорили, что старение это гетерохронный и гетерогенный процесс, исходящий одновременно из разных тканей и уровней с полиморфным проявлением. Большинство научных данных приводит к убеждению, что поскольку эти все процессы запускаются через механизмы внутриклеточного провоспаления, которое сцеплено с длиной теломер и активностью работы фермента теломеразы, которая ведет к репарациям теломер. Таким образом, все методы связанные с остановкой укорочения теломер и активированием работы теломеразы является наиболее адресным направлением по сдерживанию старения у самых различных типов тканей и при самых различных формах проявления возрастных заболеваний.
Современные подходы к лечению возрастных болезней всегда направлены на их следствия, но не коренные первопричины. Например, при болезни Альцгеймера все усилия направлены на устранение неких амилоидных белков, которые проявляют эту болезнь. Но причина глубже – отмирание стареющих клеток, которое увязано с теломерной причиной проблемы. Для этого более правильно остановить неминуемые в норме провоспалительные клеточные процессы, что поможет восстановить экспрессию генов в микроглиальных клетках, которые изначально предотвращают отложения амилоидных белков.
Современная концепция механизма укорочения теломер.
Она считает, что укорочение теломер связано с неполной копировкой с помощью фермента ДНК-полимераза. Она может копировать только в одном направлении и должна всегда держаться за маленькую часть хромосомы, она не может вернуться и скопировать пропущенные нуклеотиды. Во время репликации копируется большая часть хромосомы, но не вся: маленькая часть всегда теряется. При каждом митозе она становится чуть короче на ту часть на которой держится фермент при считывании. Со временем такая теломера укорачивается до критического состояния и отключает полностью митозы клеток. В тоже время существует фермент, который помогает клетке «вернуться» и заново удлинить теломеру, называется теломеразой.
Соматические клетки не производят теломеразу.
Дело в том, что нужный ген для этого заблокирован. Исключением из правил являются тотипотентные стволовые и раковые клетки: они производят теломеразу, которая поддерживает длину теломер, несмотря на постоянное деление клеток. Это в свою очередь противоречит общепринятой концепции о неполном считывании теломеры или отключением теломеразы.
У растительных клеток теломераза никогда не отключается.
Связано это с отсутствием у них внутриклеточной автономии по регулировке онтогенеза. В растительном мире эти права переданы с верхнего универсального уровня на уровень не просто клеток, а на многие уровни различных тканей одновременно – тканевой плюрализм. Это является огромной помехой для стволовых клеток с тотипотентным статусом, да и онкологическим. Чтобы снять противоречия с плюралистическими потребностями регулировок, изначально происходит усечение этих программ у соматических клеток.
Скорость экспрессии генов и теломеры.
Современная концепция трактует так: у молодых клеток – «молодой» паттерн = степень экспрессии генов, а у старых клеток – «старый» паттерн. Завязано это с реципрокным укорочением теломер, что и замедляет скорость экспрессии генов. На этом фоне замедляется потенциал репликации ДНК и восстановления молекул, что в свою очередь сказывается на ослаблении синтеза других молекул, совокупность действия которых обеспечивает требуемый коридор гормезиса необходимый для здоровья и молодости клеток. Постепенно клетки одновременно теряют свои функции и способность к делению. Это и есть первичный уровень клеточного старения.
Клетки с теломеразой могут поддерживать себя вечно. В них не возможно сработать триггерному механизму провоспаления. Жизненная сила клеток = Витаукт без теломеразы всегда зациклена на триггерную программу провоспаления. Инициирует её онтофизис. Чем выше продвинута онтостадийная генетическая программная реализация, тем выше провоспалительная готовность или статус. Клетка буквально источает сигналы провоспаления = SOS. Вместо репарации и пролифирации возобладают усилия на провоспаление. постепенно слабеет: теряют репаративные возможности и пролиферативную силу. Неважно, умирают они или просто становятся неподвижными и неэффективными – результат один: ослабление функциональной работоспособности ткани и её поражение и болезнь.
Каковы последствия при укорочении теломеры?
В первую очередь при укорочении теломеры укорачиваются, страдает экспрессия генов, и клетка стареет. Клетки не стареют, потому что повреждаются: они повреждаются, потому что стареют. Старение заложено на уровне программ. Похоже с этим связаны различия обмена веществ у молодых и старых людей. У стариков обмен веществ медленнее, и обновление происходит уже недостаточно быстро.
Регулировка скорости метаболизма и репарации важнейшее свойство длины теломер.
Выше мы обсуждали, что длина теломер определяет и контролирует скорость и исключает не регулируемые митозы. Но более важным является то, что длина теломер определяет скорость метаболизма за счет силы и успешности экспрессии генов, что не обязательно может быть связано со скоростью митозов. То есть, длина теломер определяет уровень жизнедеятельности не только на делящихся клетках, но и на покоящихся. Многие функциональные клетки, например нейроны, мышцы находятся почти постоянно в состоянии интерфазы, то есть пожизненного митотического покоя.
Скорость замены молекул определяет уровень функционирования клетки. Молодые клетки заменяют молекулы быстро, и большинство молекул функционируют идеально. Когда теломеры укорачиваются, экспрессия генов меняется, и необходимые молекулы заменяются медленнее. А это путь к повреждениям. Если молекулы обновляются слишком медленно, то большинство ферментов перестают работать. Это говорит о том, что обслуживающие клетки таким образом идут по пути старения. Частично обслуживающие клетки могут переходить в разряд функциональных и здесь они уже тоже имеют укороченные теломеры, что в связи со слабой экспрессией генов = слабым «паттерном» ведёт к их ограниченным возможностям по обмену веществ и репарациям. Примером этого могут быть базальные клетки кожи, которые переходят в эпидермальный слой. Это основа старения.
Проблемы начинаются, когда скорость обновления молекул уменьшается, и повреждения постепенно накапливаются. Клетки все еще работают, но становятся неэффективными. Особенно наглядно сказывается на коже, здоровье которой зависит от возобновления коллагена, да и костей, в которых развивается остеопороз. В случае когда клетки проявляют низкую функциональную работоспособность, возрастает степень возрастных заболеваний.
Скорость экспрессии генов определяет и уровень репарации ДНК.
Ремонт ДНК имеет высший приоритет в жизнедеятельности клеток. Клеточное старение тоже связано с уровнем репарации ДНК. На паттерны экспрессии генов явно накладывается, навешивается груз онтофизисного процесса и генетических экспрессий. С каждым переходом на новую стадию онтогенеза усложняется и ослабляется работа паттерна, с каждым стадийным переходом механизм паттерна становится тяжеловеснее, менее эффективным.
Суть теломерной концепции старения по Гарбузову Г.А.
Клетки делятся, ткани органов растут, органы достигают определенных размеров, это запускает коррекцию верхних уровней регулировки онтогенеза с помощью элевационных механизмов перестройки гормонального и иных профилей, последнее переключает гены клеток, причем со своей спецификой в каждой из всех систем и тканей на новый уровень, фиксируя в них уровень достигнутого онтофизиса и необходимую для этого длину теломер. На клеточном уровне это приводит к ослаблению паттернов экспрессии клеток, ограничивая их возможности в виде ослабления пролиферативной силы, репарациям, что на внешнем уровне проявляется в виде старения и болезней старости.
Эффект Хейфлика не является ограничительным механизмом продолжительности жизни животного.
Учитывая такую многопрофильность процессов старения в организме, что подобно тому как в одном организме сосуществует сразу несколько организмов и каждый из них стареет по своим принципам, это может привести к усилению диссонансов взаимодействия тканей и огромному числу возрастных болезней, с которыми организм не сможет справится и это может быть чревато патологическим процессом изнурительного саморазрушения.
Функция эффекта Хейфлика торможение скорости пролиферации и отключения линий клеток не поддающихся регулировке.
Поэтому природой предусмотрен защитный, предохранительный механизм ограничения делений или же ограничения скорости их деления = эффект Хейфлика, не допускающий организм до этих эксцессов. В культуре ткани, где обнаружен эффект Хейфлика, сняты тормозные ограничения к митозам и скорость их увеличивается, что и приводит к отключению механизма их пролиферации с помощью счетчика скорости.
В принципе он не является прямым участником процессов старения, как это подозревают исследователи, но он сдерживает принципиальные скачки излишних митозов, возможных при длительных воспалительных процессах.
Предназначение эффекта Хейфлика отключение механизмов появления недоразвитых клеток и онкологического перерождения быстро пролиферирующих клеток.
Чрезмерные митозы усиливают внутриклеточные механизмы провоспаления, а это накаляет ситуацию, ведет к образованию неуправляемых недоразвитых «тупых» = «слепых» клеток, что является начальным этапом их онкологизации. Очевидно, такие «слепые», недозрелые клетки не успевают сформировать на своих мембранах сенсорные структуры, рецепторы и становятся чужими.
Ярким примером этого может быть белокровие = лейкоз, когда в крови плавает избыток недоразвитых белых телец. Их бы и пресечь с помощью механизма Хейфлика = накидывание на них сети, ловушки Хейфлика, но… процесс заходит слишком далеко и эффект не срабатывает, а недозрелые клетки при достижении некой критической массы приспосабливаются путем включения теломеразы – фермента присущего только генеративным половым клеткам. Этим они превращают себя в бессмертных, мутируют, закрепляют это свойство в геноме за счет митохондриальной дисфункции.
Следовательно, эффект Хейфлика, так же как и механизм апоптоза, – защита от онкологического перерождения. Ограничительные возможности длительности жизни человека и его старение не связаны с укорочением теломер. Скорее всего, этот счетчик показывает сколько раз организм заходил в провоспалительное состояние, когда потребовалось бы запускать чрезмерную пролиферацию. Он усекает работу только линий чрезмерно пролиферирующих. Он «отлавливает», ограничивает такие ситуации.
В обычной жизни такие ситуации, когда ограничение жизни связано с отключением митозов в одной из пролиферирующих линий, не доходит, не доживают. Организмы животных умирают от старения задолго до того как исчерпывается потенциал теломер.
Теломерная концепция старения.
Одна из современных концепций старения у животных увязывает её с утратой у хромосом теломерного счетчика делений к восстановлению. Это в свою очередь связано с блокировкой фермента теломераза, ответственной за восстановление теломер.
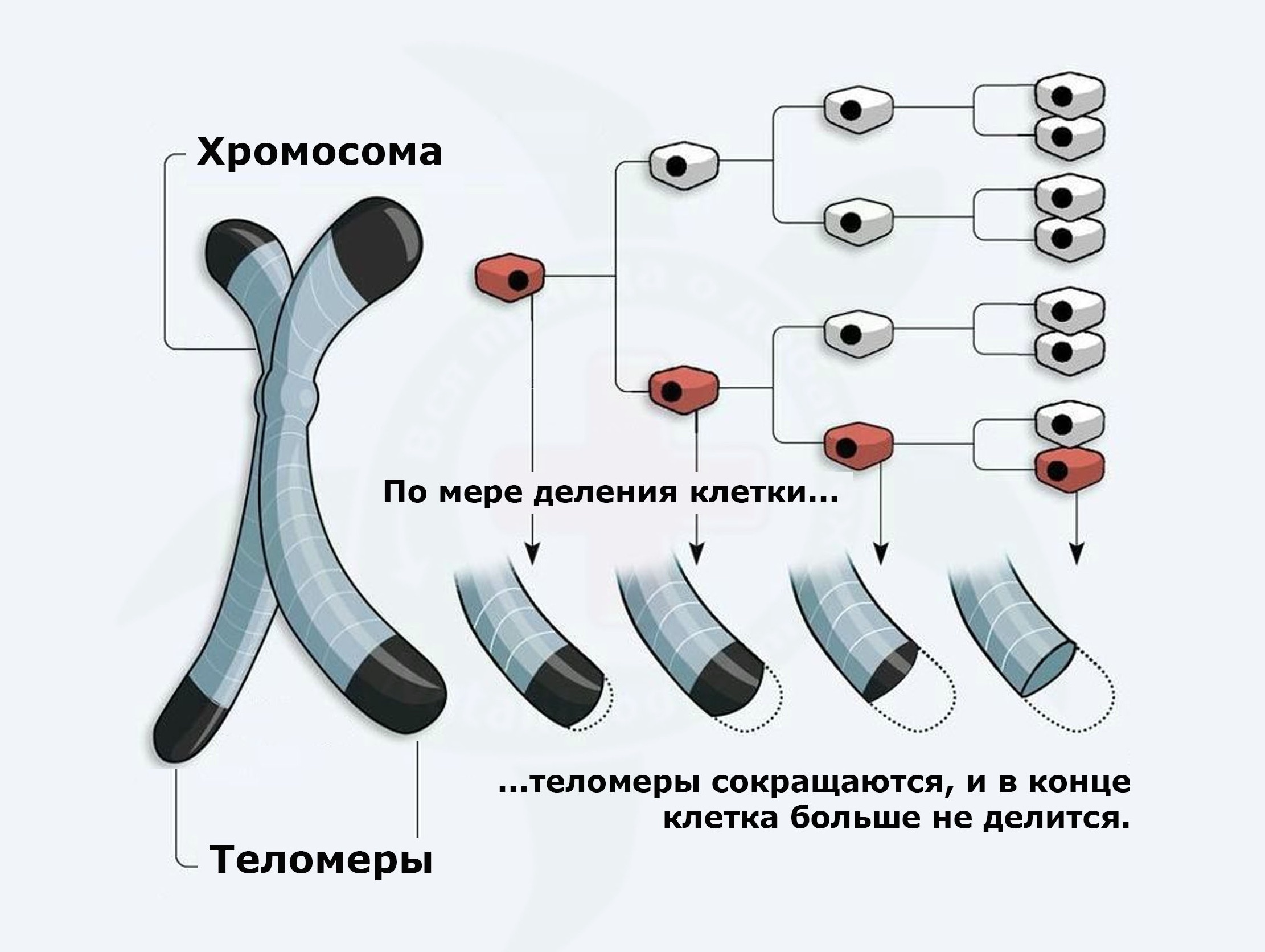
На рисунке: Схематическое изображение теломерных колпачков на кончиках хромосом, укорочение которых ведёт к остановке роста клеток.
Ведутся активные исследования по способам активизации фермента теломераза, благодаря которому могло бы происходить постоянное обновление укороченных теломер. Господствующая концепция гласит, что восстановление теломер позволит стволовым клеткам возвращать их к юношескому состоянию. В основе действия таких препаратов лежит воздействие на теломеры, которые защищают ДНК от повреждений.
При введении теломеразы клетки фибропластов в пробирке вместо 80 раз делились 280 раз. Похоже этот эксперимент ничего не доказывает в плане омоложения, но ведёт к извращенной реакции по пресечению появления недоразвитых клеток.
Изучается и механизм управления этим ферментом. Это якобы позволит приостановить процесс старения и массовой гибели клеток.
Исходя из такого ракурса в пересмотре бывшей концепции следует, что ограничительные механизмы и старение начинаются не на уровне клеток, а на уровне межклеточного взаимодействия. Одни ткани побуждают к старению другие.
В то же время появились данные утверждающие, что восстановление теломер якобы позволяет продлить жизнь подопытных животных. Так ли это? Ведь по нашей концепции снятие теломерной ловушки должно усилить появление недозрелых клеток, а значит болезней возраста.
Омоложение тканей мозга – должно иметь лидирующее значение.
Считается, что старение организма человека начинается с мозга, который инициирует этот процесс. Выдвинуто предположение о возможности образования у пожилых животных новых нейронных связей в гиппокампе - области, жизненно важной для памяти и особенно чувствительной к процессу старения.
В этих экспериментах в качестве донорского материала использовали пуповинную = плацентарную кровь человека: цель изучить белки плазмы, которые являются общими для мышей и людей и чей уровень снижается с возрастом.
Найдены два претендента на звание самого главного белка, контролирующего молодость мозга - CSF2 и TIMP2. Один из них связан с накоплением токсичных белков и различными нарушениями когнитивных функций. Роль второго белка тоже связана с молодостью мозга: при искусственном удалении TIMP2 из крови мышат, у них резко падала острота памяти и нарастали симптомы старческой деменции. Этот белок служит регулятором генов, участвующих в росте и развитии клеток. И его силу, а также силу других регуляторных компонентов молодой крови, можно использовать для коррекции возрастных нейродегенеративных заболеваний у людей.
Это повышало регенеративные возможности многих тканей, в том числе поврежденных тканях печени, сердца, мышц, костном мозге, улучшались когнитивные способности: память, внимание, ориентирование. В свою очередь факторы старой крови вызывали обратный эффект в виде торможения процесса образования новых клеток. Эффект сенелитирования = состаривания был выше, чем эффект реювенилизации. То есть старая кровь сильнее чем молодая. Легче сенелитировать организм чем его реювенилиировать. Похоже, что в стареющих клетках существует избыток неких белков и молекул, уровень которых надо скорректировать, прежде чем ожидать эффекта от молодой крови.
С другой стороны важен вопрос: удастся ли с помощью такого стадийного омоложения добиться и сдерживания старения на клеточном уровне? Интуиция подсказывает, что это возможно только частично, так как это самостоятельное направление, имеющее иные механизмы старения и на другом уровне. Клеточное старение это всего лишь первичный элемент стадийного старения, когда включены программы на ограничение на этапе раннего эмбриогенеза. В самом начале становления особи появились программы ограничивающие её возможности, первые заложенные программы проявления её старения. Следовательно, работая с коррекцией гормонального профиля, мы не сможем достичь коррекции на более глубинных первичных механизмах, запрограммированных изначально процессов на уровне клеток, связанных с первичными факторами роста, вернее прото- факторами = древними = изначальными, которые вырабатываются только на этапе эмбриогенеза. Эти программы на полное самовосстановление отрезаны изначально на самом раннем этапе = зиготном становления организма.
Клеточное независимое старение у животных можно обозначить как одну из стадий онтогенеза, когда на уровне эмбриогенеза закрыты программы на полную реювенилизацию и тотальную регенерацию.
Похоже важной чертой клеточного пути старения является снижение регенеративных свойств тканей организма, обусловленное нарушением функций стволовых клеток. Стволовые клетки здесь изначально не имеют той силы, которая возможна в эмбриогенезе. Конечно, регенеративную способность может ограничить и особенности гормонального фона и адальтус с сенелитом. Но наибольшая часть возможностей утрачивается не в связи с гормональным профилем, а с первичными задатками, потенции к этому.
Межтканное взаимодействие может определять скорость старения одной ткани на другую.
Существуют белки недостаток которых в красных кровяных тельцах ведет к снижению когнитивных способностей и преждевременному старению. Следовательно старение ткани может инициироваться не только в пределах данного органа или системы, но и извне.
Выяснили, что снижение в крови уровня белка ADORA2B ведет к ухудшению слуха и памяти, начинается воспаление в головном мозге, и старение наступает быстрее. Это так называемый "антивозрастной белок".
Красные кровяные тельца выполняют функцию доставки кислорода в клетки тела. Сигнальный каскад красных кровяных телец ADORA2B, способствуя доставке кислорода в мозг, борется с ранним началом возрастного снижения когнитивных способностей, ухудшением памяти и слуха.
Молодая кровь против болезни Альцгеймера.
Существует экспериментальный проект по лечению этой болезни с помощью переливания крови. Одни больные в течение месяца получали плазму от 18-30 летних доноров, а другие - инъекции плацебо. Оказалось, что вливание свежей крови успешно сказалось только на рутинных навыках людей без видимого улучшения общих когнитивных качеств. Несмотря на неоднозначные результаты экспериментов, исследователи не отчаиваются и настаивают на продвижении этого метода.
Возможности омоложения вилочковой железы.
Особенность тимуса в том, что с возрастом он исчезает. Причём исчезать начинает уже во время полового созревания, к старости же этот процесс особенно ускоряется.
Таким образом, это одно из многочисленных самостоятельных проявлений онтостадийных переформатировок, перестроек, но не является следствием старения.
В эксперименте использовали стволовые клетки, которые активировали так, чтобы они возместили возрастной ущерб с помощью определенных белков, которые переключали нужные гены. Стимуляция их у старых мышей приводило к тому, что переродившийся и уменьшившийся с возрастом тимус у старых мышей заметно увеличивался. Причём не просто увеличивался, но и начинал производить больше клеток. То есть его функция тоже восстанавливалась. Введение пептидных белков этой железы увеличивало компетентность иммунных клеток в старом организме и продолжительность жизни животных.
Иными словами, удалось заставить орган регенерировать прямо в живом организме!
В частности, нужно понять, насколько функциональны Т-клетки, которые производит возрождённый тимус, и действительно ли такая регенерация улучшает защитные свойства иммунитета.
Похоже это пример гетерохронности старения среди разных органов и возможные первичные их механизмы могут быть как верхними, так и на нижних этажах.
Саркопения = потеря мышечной массы – элемент старения.

На фото: Выраженный пример саркопении как одного из элементов старения.
В ряде случаев можно наблюдать преобладание ускоренного старения мышц по сравнению с другими тканями. Но чаще они равномерно стареют с другими тканями, что проявляется в их ослабевании, заметного уменьшения массы – это характерный признак.
Саркопения так же неотвратима, как и старость. По каким-то внутриклеточным причинам клетки мышц перерождаются и начинается атрофия мускулатуры.
Остеопения = остеопороз – одно из проявлений старения в костной системе.
Проблема, которая проявляется как неминуемый компонент старения в костной системе. В видении некоторых ученых в будущем даже люди в возрасте 85 лет и старше смогут вести активную жизнь без риска переломов, развития рака и других проблем старости.
Такие надежды связаны с белком RANKL, который является главным регулятором процесса дегенерации костной ткани. Разработан препарат, который приемлем для лечения остеопороза и ассоциированных с раком патологий скелета.
Кроме того, установлено, что этот же белок вовлечен в регулируемый половыми гормонами механизм запуска лактации при беременности. Одновременно показано, что действие этого белка сопряжено с гормональным профилем. Это позволяет увидеть начало цепочки старения на клеточном уровне в частности в костной системе с более общим процессом предшествующих изменений гормонального профиля.
Пересадка органов и онтофизис.
Известен случай пересадки печени молодого человека 20 лет старому, что привело к явным процессам глубокого омоложения, вплоть до восстановления седых волос в черные, улучшение зрения и многих иных внешне хорошо проявляемых признаков. Следовательно и у человека стадийный сенелит это не необратимый процесс. Реювенилизация онтостадийно зрелых и старых особей возможна и в мире животных.
Реювенилизация у онтофизисных привоев плюща оказалось связана с особо высоким содержанием ауксинов у подвойных ювенильных растений плюща. Обработка взрослых растений плюща ауксинами позволяла им начать образовывать корни и изменять форму листьев, что говорит о возврате к ювенильному состоянию. Даже если в один сосуд поместить вместе ювенильные и взрослые растения плюща, то у взрослых образуются корни, как и при обработке ауксинами. При этом их образование сопровождается возвратом к ювенильной форме листьев и задержкой зацветания.
Возможности омоложения кроветворной системы.
Доказана возможность функционального омоложения гемопоэтических, то есть стволовых клеток, что соответствует нашей концепции гетерохронности и гетерогенности, то есть двойственности механизмов старения, исходящих как с нижнего клеточного уровня, причем из различных тканей по своим различным руслам, так и с верхнего, элевационного процесса на уровне изменения чувствительности гипоталамуса с переходом на новый гормональный профиль.
Стареющие кроветворные клетки утрачивают свою структурную организацию и полярность, что приводит к снижению их функциональной эффективности. Найдены ингибиторные белки, которые тормозят первичные клеточные механизмы старения, что улучшало состояние и восстанавливало функциональность до уровней, характерных для молодых клеток.
Иммунопения и возможности омоложение иммунитета.
Подтверждена возможность воздействия изолированно только на эту систему, что никаким образом не влияет на другие системы. Это подтверждает, что каждая система имеет свои самостоятельные зачатки старения.
Иммунопения = процесс старения иммунной системы. Многочисленные исследования показали, что с возрастом выработка B-клеток сокращается. Кроме того, начинаются сбои в процессе отбора, в результате чего накапливаются старые клетки с ограниченными возможностями.
Для этой системы найден способ обратить вспять этот процесс путём удаления старых В-лимфоцитов из организма пожилых мышей и поощрения выработки молодых и мощных клеток. Подобно всему организму, иммунная система ослабляется с возрастом, что приводит к существенному росту заболеваемости среди пожилых людей и резкому снижению их способности реагировать на вакцинацию. В-лимфоциты являются основными клеточными компонентами иммунной системы и несут ответственность за производство антител.
Оказывается, для восстановления B-лимфоцитного гомеостаза достаточно искусственным образом вызвать хронический дефицит этих клеток. Организм вынужден вновь активировать костный мозг и заставить его производить B-клетки в объёме, который не отличается от показателей молодых мышей. В результате ответ на вакцинацию у пожилых улучшался в 5 раз. Удалось доказать, что физиологическое старение регулируется процессом, который можно отменить.
Восстановление стареющего иммунитета.
С увеличением продолжительности жизни всё сильнее вскрываются элементы старения иммунитета, которые на других стадиях онтогенеза малозаметны. Одной из находок ученых была идентификация молекулярного «переключателя», позволяющего активировать иммунные клетки, известные как натуральные киллеры. В экспериментах на мышах показано, что белок Cbl-b (известный также как протоонкоген Cbl) блокирует способность натуральных киллеров распознавать и уничтожать злокачественные клетки.
Инактивация этого белка восстанавливает дееспособность натуральных киллеров, что в свою очередь позволяет им более эффективно выполнять свои функции и активно подавляют метастазирование опухолей. Иммунная система нужна для того, чтобы выявлять опасности и реагировать на них. В норме предшествующим всему этому является механизм провоспаления. В то же время при старении в организме формируется свой так называемый воспалительный статус, который мешает нормальному проявлению первого, что и сказывается отрицательно на состоянии здоровья и мешающий иммунной системе эффективно распознавать и уничтожать злокачественные клетки.
Полученные результаты позволят начать разработки нового поколения препаратов, которые помогут иммунной системе лучше справляться с осложнениями процесса старения и позволят человеку жить долго и умирать без многолетних страданий, причиняемых хроническими возрастными заболеваниями.
Его определяет возрастная динамика гормонального профиля, которая в свою очередь определяет действие белка RANKL, что является ключевым инициатором развития гормонозависимого рака молочной железы. Это пролило свет на молекулярную взаимосвязь между заместительной гормональной терапией, приёмом гормональных контрацептивов и раком молочной железы. Из этих данных видно, что верхний уровень изменений подстраивает под себя нижний уровень настроек и тем самым определяет возможность нижнего клеточного уровня проблем. Наступление нижнего уровня проблем невозможно без инициирующего действия сверху.
С другой стороны на проблему можно посмотреть со стороны клеточного уровня и предположить, что это связано с первичным изменением чувствительности гормонозависимых тканей и их клеток к гормональной регулировке. Но гормональная кастрация может отменить наступление этой проблемы, что означает, что первичные корни здесь на верхнем уровне регулировок.
Итак, существует ось регулировок: верхние программы, которые обеспечивают динамику гормонального профиля = элевационные механизмы, и нижний клеточный уровень геномно-эпигеномных подстроек, приводящий его в соответствие с верхним. Каждый из них имеет свои собственные механизмы и генопрограммы.
Постэмбриональный онтофизис - как причина утраты способности к полной реювенилизации на клеточном уровнем = плацдарма для старения на клеточном уровне?
Создается впечатление, что это независимый от стадийного старения процесс? Клеточный уровень старения у животных похоже связан с утратой ими эмбриональных потенций, наиболее первичных этапов самоомоложения и регенерации. Клетки животных и их стволовые линии сохраняют постэмбриональное состояние онтофизиса и поэтому не могут полноценно омолаживаться. Программы полноценного самовосстановления у них усечены еще на стадии зиготы. Это и объясняет ограничительный эффект Хейфлика.
Различные типы старения связаны с гетерохронностью и полиморфизмом темпов старения различных тканей.
Хорошо известно, что большинство людей стареют по разному и длительность жизни разная. И в тоже время в них есть определённая общая доля закономерности. Поэтому применяют понятия биологический возраст и паспортный. С этим же связано и понятие преждевременное старение. Его определяют как совокупность скорости старения разных органов и систем организма, а также связывают с генетическими предрасположенностями.
Метилирование ДНК - один из универсальных процессов старения на клеточном уровне и являющийся генетическим показателем старения во всех тканях.
Оно определяет биологический возраст человека. Это эпигенетический, то есть не затрагивающий саму последовательность ДНК, процесс. Суть его в изменении "надстроек" на молекуле ДНК, которые приводят к экспрессии или репрессии определённых программ. Эти надстройки похоже несут функции квазипрограмм и помогают, подготавливают геном к блоковым «включениям» = экспрессиям, то есть включения целой сети сопряженных программ. Очевидно, на этих рельсах зиждется вторая сторона оси регулировок онтогенеза, не связанная с верхними механизмами регулировок, то есть с динамикой гормонального профиля, определяющая свой верхний комплекс программ онтогенеза, но связанная с комплексом возрастных генопереключений, которые снизу определяют и закрепляют их уровень онтостадийности.
Итак, эпигенетические изменения приводят к изменению работы ДНК, без изменения генетического кода. Устоялась концепция, что метилирование ДНК – это процесс, который происходит равномерно и постоянно в течение жизни. Однако другие учёные выяснили, что метилирование ДНК в организме каждого конкретного человека происходит неравномерно.
Выяснилось, что индивидуальные "траектории" метилирования ДНК у всех этих людей отличались в зависимости от целого ряда показателей, таких как генетика и окружающая среда. Это помогло разработать новый метод определения биологического возраста человека на основе особенностей метилирования ДНК, которые определяют "индивидуальные треки старения". Люди действительно стареют по-разному. Данные показывают на неоднородность в метилировании ДНК, которую наблюдают у пожилых людей и людей с врождёнными и возрастными заболеваниями.
Проявления онтостадийных различий в одних и тех же тканях у молодых и старых животных.
Важным проявлением отличия заключается в том, что у старых особей намного выше провоспалительный статус и наиболее узкий коридор потенциальных возможностей противодействия нагрузкам, то есть гормезиса. Степень провоспаления сопряжена с уровнем аминокислоты триптофана. Так молодые грызуны при низком уровне аминокислоты не испытывали никаких симптомов, а у старых мышей при недостатке триптофана возникло воспаление кишечника. То есть склонность к провоспалениям у них намного выше и это связано с онтофизисными перестройками.
Итак, повышенная готовность или статус провоспаления – основной признак старения, но важнейший билогический вопрос: откуда исходят его первичные корни - из клеточного уровня или более верхнего уровня? Похоже это сугубо онтостадийное последствие перестроек, на нижнем уровне, но являющееся следствием перестроек на верхних уровнях. Любая автономная система – это саморегулирующаяся независимая система. В описываемом нами случае в молодости эта система не чувствительна к недостатку метионина и работает на принципах оптимума коридора гормезиса, у неё есть запас прочности. Но та же система с возрастом проявляет другие свойства в виде провоспаления на любые перекосы в метаболизме, так как запас прочности здесь минимален и связан он с внутриклеточными геномными перестройками. Но автономия здесь не полная, а строго на определенные параметры требуемые необходимостью данной квазисистемы регулирования в данном органе или ткани. Часть функций управления остаётся на верхнем «федеральном» уровне. Каждый из управленцев решает свой круг вопросов.
У растений права на автономию даны только в вопросах морфогенеза, роста, тогда как вопросы онтогенеза полностью подключены через федеральный = центральный уровень. Да, у растений есть и клеточно-тканый уровень регулировки и закрепления стадийности онтогенеза на нижнем уровне в виде топофизиса и циклофизиса, но здесь их запускают верхние процессы федерального уровня. В механизмах регулировок у животных больше прав на автономию и они своими местными механизмами регулировок задают план действий центру. Кроме того, здесь имеются «республиканские» уровни регулировок, так как здесь на много больше систем и органов работающих по своим правилам развития и требующих пере подключение регулировок на себя.
У животных всё намного сложнее: здесь существует единая ось с привилегией регулировок на нижнем местном уровне.
Содержит ли сенелит в себе мортал-фазу = элемент смертности?
Все стадии онтогенеза имеют в себе программы продвижения в другие стадии, кроме сенелита, где таких программ больше нет. В этой стадии организм теряет способность к восстановлению. Здесь особенности скорости метаболизма и репарации настолько снижены в соответствии с сенелитной программой записанной на генетическом уровне, само по себе является уже стрессом, но уже непреодолимым.
Итак, создается такая генетическая обусловленность, когда скорость восстановления как от стресса, так и по внутренним особенностям сенелитного состояния уменьшается вдвое, что и является возрастным ограничителем. Чтобы предотвратить мортальный исход, необходимо не доводить вход организма в сенелит, так как последний содержит в себе такую скорость метаболизма, которая зафиксирована генетически через активность работы митохондрий, и не совместима с параметрами ВИТАУКТА и ГОРМЕЗИСА.
В возрасте 120-150 лет организм человека полностью теряет способность к восстановлению, а значит любой стресс, патогенетический фактор может оказаться фатальным.
Причины ограничений = клеточного старения = лимитов, которые исходят из клеточного уровня у животных.
В растительном мире отсутствуют клеточный уровень лимитов и старение.
Анализ всех возможных способов старения в мире растений показывает, что здесь нет старения исходящего из потенциального ограничения возможностей на клеточном уровне. А это означает, что нет оксидативного стресса, механизмов провоспаления исходящего из старения клеточных линий, то есть на клеточном уровне и эффекта предела митозов Хейфлика, нет митохондриальной возрастной недостаточности и теломеразного ограничительного счетчика, а также нет онкологизации. Даже у короткоживущих 1- и 2-летних растений старение особей запускается на верхнем общеорганизменном уровне, то есть путём гормонального контроля. Почему такие большие отличия с миром животных? Возможно, это связано с делегированием = передачей части функций = полномочий регулировок с верхнего организменного уровня на нижний клеточный уровень? Или это связано с эпигенетическим уровнем наподобие тому, что нами описано выше как топофизис ил циклофизис у растений, то есть закрепление в клоновых потомствах некоторых приобретённых с возрастом особенностей растения?
После завершения этапа имаго = юности размеры организма полностью прекращают увеличиваться, количественный рост замещается на качественные изменения связанные с созреванием. Происходит переход регулировок с общеорганизменного уровня, когда регулировка шла от внешних параметров, на нижний клеточный уровень регулировок. Это более тонкая и вынужденная особенностями жизни животных регулировка.
Особенности регулировок роста у организмов растительного мира.
У растений как морфология, так и онтостадийная зрелость и рост определяются преимущественно сверху, «централизованно» – гормональным профилем, но не выходя за рамки программ генетического контроля. В животном мире дано больше прав «автономии», на месте.
Здесь осуществляется замкнутый круг прямых и обратных связей: верхний гормональный уровень регулировок определяет контроль за генетическими программами, их переключение, а последние определяют рамки возможностей, коридор, по которым может пойти развитие во внешнем проявлении. Всё осуществляется, «едет по рельсам», которые обеспечивают гено-программы, но реализуют их гормоны.
Уровень из которого исходит начало старения: из центра или автономии?
Ответить, что сверху или снизу – будет неправильно, так все они закольцованы и выступают как одно единое целое. Это два рычага, которые у одних видов могут больше реализовываться с нижних этажей, а в других – с верхних. Всё зависит от видовых потребностей. Если бы не было ограничивающих моментов, то рост мог бы происходить неограниченно. В то же время в пределах одной особи этого достичь невозможно, так как рост такого растения-дерева сам себя ограничивает как в ширину, так и в высоту. Растение исчерпав возможности вегетативного неограниченного роста, например у бамбуков, переключает механизмы перехода из вегетативной фазы в генеративную. В свою очередь растения с фиксированными сроками плодоношения изнутри получают такой сигнал. Там, где начинают включаться механизмы ограничения роста = ювенила, там запускаются механизмы адальтуса, а затем матуры и сенелита. Включающие механизмы могут ориентироваться на внешние или внутренние факторы.
Сенелит может быть как запрограммированным организованным процессом на самоисключение, самоотключение, так и не запрограммированным и происходить произвольно, естественным путём, когда особь выходит из оптимального коридора существования и внешние факторы становятся сильнее для растения и добивают его постепенно. Внутренних программ противостояния у растения этим факторам больше нет, они исчерпаны. Жизненные силы становятся слабее внешних факторов. Поэтому старение не обязательно проявляется в виде программы, часто оно является следствием отсутствия программы. Так медленно стоя, отмирают деревья гиганты.
Во всех случаях внешней или изнутри исходящей регулировок общим является, что все они регулируются через онтостадийные механизмы.
Конечно, старение завязано с хронологией жизни растения, его этапами. Скорее всего правильнее ответить – это динамический процесс, который раскручивается как снизу, так и сверху, когда идёт движение по ступеням реализации комплекса программ развития, но на определенном этапе хронологии входит в зону неоптимального существования, когда оно может быть обусловлено как изнутри генетически, так и внешними факторами.
Особенность животного мира - появление генетического происхождения онкологических болезней.
Растительный мир свободен от мутационного происхождения онкологии. Здесь если и есть опухолевые разрастания, то они все связаны с внешней паразитической инвазией, а сугубо сопряженных с клеточными мутациями не бывает. Это удивительно, но факт. В тоже время в человеческой популяции каждый второй умирает от рака. Трактовать это можно тем, что на программы промоуции = делений «навешено» множество и других ограничительных программ регулировок роста, исходящих изнутри самих клеток, а не из общих программ развития.
Множество «приставных» программ связано с фиксированным ростом у животных. Каждый вид ткани имеет свои ограничения и свои специфичные механизмы регулировки роста. Причем, процессы образования новых клеток жестко сбалансированы с отмиранием и апоптозом клеток. Рост регулируется процессами отмирания. Сколько клеток отомрёт, столько 1:1 должно нарасти. Такая точная регулировка возможно только на уровне взаимодействия клеток данной ткани = межклеточное взаимодействие, что не достижимо на уровне организма.
Рецепторные механизмы регулировок – особенность присущая животному миру.
Для межклеточного взаимодействия природа придумала рецепторные механизмы чувствительности, чтобы данный сигнал, регулировка шла только адресно извне или в пределах данной ткани. Все мембраны клеток это огромное сложное сенсорное поле, которое должно получать огромное количество корректирующей информации. Такого сложного уровня регулировок в растительном мире и близко нет. Если у растений механизмы регулировки роста тканей, «включатели» роста осуществляется на общеорганизменном уровне и ориентируется относительно верхних параметров, например гормонального профиля и иных параметров, то у животных на первый план по значимости выходит внутритканный уровень регулировок. Каждая ткань сама заботится о своих величинах и темпах клеточного роста. Для этого ей необходимы свои особые механизмы, которых нет у растений.
Условно растение можно представить как единый сложный орган управляемый общей системой регулировок и интегрированных многочисленных функций, тогда как организм животного это совокупность систем, это многоэтажная интеграция интегрированных органов. То есть это сверхинтеграция, механизмы управления в которой намного сложнее. Каждый орган (их 78) и система (их 12) здесь накладывает свои ограничительные требования и всё это многообразие требований необходимо записать по отдельности в генетическом коде, а затем согласовать всю эту сложную иерархическую совокупность с помощью генетических блоков и включателей. Вся эта сложная конструкция должна находиться в гармонии и развиваться по общим единым правилам. Но эта же конструкция придаёт такому организму огромное количество ограничений вплоть до лимитирования его продолжительности жизни и огромному количеству возможных заболеваний, например вплоть до повышения вероятности неминуемой случайности проявления онкозаболеваний до 50% с 0% вероятностью у растений. Это болезнь сугубо животного мира.
Очевидно, здесь кроются причины появления возможности в животном мире онкологических проблем, то есть когда «нежные = тонко настроенные» генетические программы не справляются с противодействием нагрузкам, которые обеспечивают заданные гомеостазы.
Слом сенсорных структур на мембранах клеток – начало к онкологическому перерождению клеток.
В отличие от растительного мира в животном мире хорошо развита сенсорная = мембранная система регулировок. Эти механизмы обеспечивают значительно повышение статуса и возможностей независимости клеточного уровня регулировок.
Этому способствуют в первую очередь нагрузки на сенсорные устройства обеспечивающие их. Сенсоры и рецепторы это сложные конформационные белки – объемно-пространственные или же работающие по принципу: ключ-замок. От их регулировки зависит работа митохондрий. В ряде случаев возникают сверхвысокие потребности в кислородного типа, то есть митохондриальной энергетике, необходимой для сохранения дифференциальной работоспособности. В первую очередь при онкологии выгорают цилии = антенны и рецепторы. Это приводит к тому, что клетка становится «глухой» не восприимчивой к сигналам и начинает работать вразнос её энергетика с переходом на аэробные гликолизные типы энергетики. К этому всему подключаются механизмы провоспаления, которые могут проявить себя двойственно и повести ситуацию в положительную или негативную сторону регулировок. Как это случится управляют более глобальные механизмы регулировок на уровне программ гормезиса, отвечающих за подключение и слаженную работу одновременно множества программ. Здесь уже очевидно вмешивается и уровень квазипрограмм и эпигеномного управления, последний из которых зависит от общего здоровья и состояния данных клеток. Онкологизация возможна сугубо как на этом клеточном уровне нарушения регулировок, так и особо на фоне онтостадийных клеточных и общих перестроек.
Неуправляемость клетки – причина работы вразнос митохондриальной кислородной энергетики.
Это и есть прямой путь к онкологизации = онкологическому перерождению клетки. Аэробная активность митохондрий регулируемый сенсорными структурами процесс. При «выгорании» цилий происходит нарушение регулировок кислородного метаболизма и он переходит частично на гликолизный тип, анаэробный метаболизм. Отсутствие цилий ведет к отсутствию тормозов, тонкой настройке. Запускаются механизмы оксидативного стресса и провоспаления = тлеющего пожара. В норме последние в свою очередь должны запустить механизмы апоптоза или некроза больных клеток. Но этому мешает предшествующая этому десенсибилизация клеток, что делает её неуправляемой, нерегулируемой. Начинается противоречие, несогласование программ. Одна из программ становится лишней. Здесь кроется причина их «поломки» = мутации. Оксидативный стресс, избыточные радикалы ведут к перегреву «сжиганию» лишних программ. Напряжение на эти программы неминуемо вызывает подключение защитных внутриклеточных механизмов провоспаления, а затем и работу вразнос энергетических станций – митохондрий. В таких особо «горячих» условиях, точках напряжения особо легко прорвать, сорвать работу программ, такие программы сначала «зависают», а затем портятся. Происходит слом программы регулировки роста в конкретном месте в конкретной ткани, например коже. В каждом типе ткани происходит образование своего типа опухолевого процесса, то есть нерегулируемого роста клеток, присущего данной ткани. Локальные, региональные механизмы регулировок, торможения клеточных делений становятся на пути более общих механизмов регулировок. Клеткам становится тесно в этих ограничительных промоуторных рамках. Более древние прото-программы прорывают более поздние прима-программы.
Каждому типу ткани присущ свой тип опухоли, а значит и специфические её особенности. Усложнение механизмов клеточных регулировок в животном мире приводит к появлению онкологических проблем.
Особенности старения в животном мире связаны с подключением клеточных лимитов и старения.
У организмов животных сенелит регулируется не только через онтостадийные механизмы, но и на нижнем уровне: клеточном и межклеточного взаимодействия. С этим же связана блокировка программ большой или тотальной регенерации. Эти программы находятся в антагонизме, противоречии с программами межклеточной регулировки. Поэтому более общие общеорганизменные программы более глубоко запрятаны = заблокированы. Легко обслуживаемыми, доступными программами остаются только межклеточных взаимодействий.
Допустим где-то локально в организме поврежден какой-то орган или ткань, то это не значит, что для локального восстановления нужно восстанавливать всё, как это возможно в мире растений. Нужна только локальная регенерация и то в пределах только размеров той ткани, в которой произошло повреждение. Общеорганизменный уровень регулировки здесь не нужен и даже вреден. А это можно предусмотреть только в пределах межклеточных взаимодействий.
Механизмы межклеточных взаимодействий и регулировок.
Эти программы сцеплены не с уровнем гормональных регулировок, а с клеточными потенциями, которые уже управляются не из вне, а из изнутри самой данной автономной единицы: системы или ткани. Для этого могут быть задействованы белковые факторы роста, специфичные для каждой ткани или системы. Существует 12 систем организма, которые обслуживают конкретный специфичный для них ряд тканей. Регулировка здесь осуществляется через сигнальные молекулы, действующие на рецепторные структуры сенсорного дисплея мембран или же непосредственно на гены хромосом.
Механизм внутриклеточных регулировок.
Кроме уровня межклеточной регулировки, очевидно, существует и уровень сугубо клеточных потенций, независимо от того какие они ткани представляют. Это самый первичный уровень, который задействует гено-ключи как тоти-, плюри- …, то есть эмбрионального развития. Этот уровень менее всего поддаётся внешним коррекциям. Он как бы изначально задан, хотя это тоже программа, но самая первичная.
У взрослых организмов эти ключи от первичных «комнат развития» не производятся, а ограничены только уни- и монопотентными ключами, что позволяет обеспечить только малые, локальные, на уровне данной ткани регенерации. У большинства животных механизмы регулировок этих стадий развития запрятаны на уровне геномов клеток, когда они отвечают только за эмбриональный рост и развитие. Это говорит о том, что пробуждению этих программ внутри клеток мешает экспрессия каких-то надпрограмм или блоки внутри клеток. Открыть первые означает закрыть вторые, что возможно поведет к необходимости дедифференциации. На это способны некоторые гено-ключи. Так например, разблокировка этих клеточных генопрограмм частично реализуется у аксолотлей, когда регенерируют полностью отсеченные конечности.
Похоже, в животном мире эти программы полной или большой регенерации вредны. Например, они могут противоречить программам стадии матуры. Это два противоположно действующих процесса и взаимоисключающих друг друга? Поэтому те возможности к пролонгированию роста и омоложения, которые имеются в растительном мире, практически исключены в животном мире, запрятаны в геномный уровень регулировок.
Важным условием особой = на клеточном уровне регулировки онтогенеза у животных является наличие механизмов фиксированного роста и размеров.
Особенностью животных являются наличие обязательных механизмов, сохраняющих пропорции в размерах тканей и органов. У растений этих лимитов нет. Фиксирование размеров и роста у животных компенсируется удлинением фаз адальтуса и матуры = созревания и перезревания. Высокая степень матуры полностью перекрывает = блокирует программы на регенерацию и омоложение = ювенилизацию и ревитализацию. Чтобы вернуть возможность включать механизмы самоомоложения, необходимо предварительно устранить матуру за счет возврата к неотении.
Таким образом, в животном мире можно предполагать имеется три уровня старения:
- это верхний, который управляется онтостадийным развитием и обеспечивается гормональным профилем;
- это средний = межклеточный; обеспечивающий фиксированный рост, а также это уровень органов-мишеней, который может управляться и сверху, и снизу;
- это снизу, исходящий из клеточного уровня, ограничения здесь связаны с блокировкой программ на этапе эмбриональной диапаузы, то есть на эмбриональной фазе развития.
Программы связанные с тотальной возможностью к регенерации = тотипотентные здесь запрятаны = заблокированы на уровне эмбрионального развития, когда только и возможно производство тотипотентных гено-ключей. У высших животных полная регенерация и омоложение неосуществимы уже на уровне первичных эмбриоблоков. С этим же связаны ограничения механизмам в противодействии старению. У человека большая часть проявлений старения исходит не из стадийных верхних уровней процессов сенелита, а из клеточного уровня.
В тоже время взлом этих программ эмбриональных блоков открывает брешь к онкологическим процессам = неограниченному неуправляемому росту. Ломать нельзя, нужна лишь управляемая подконтрольная корректировка. Вмешиваться в эти эмбриопрограммы можно, ведь природа это делает повсеместно легко и просто, но методами правильного балансируемого управления. Плющ например из стадийного зрелого плодоносящего состояния легко переходит при измене среды в ювенильную фазу, у аксолотлей отрастают новые конечности.
Клеточный уровень старения всегда запускает провоспалительные механизмы сами в себе.
Следует заметить, что старение всегда является и одновременно исходящим из клеточного уровня провоспалительным процессом. Нет, иммунного уровня механизмов воспаления здесь нет. Сигнальные вещества на воспаление выделяют сами стареющие линии клеток. Такие клетки не могут поддерживать высокий уровень дифференциации? Высокую степень дифференциации им обеспечивает особо высокий уровень, накал работоспособности митохондрий. На постоянном пределе возможностей работают, например, нервные и гормональные клетки. Они медленно деградируют и перерождаются = дегенерируют в низко специализированные, кислородомалозависимые фиброзные клетки. Им не комфортно существовать в условиях ограниченного сдержанного роста, связанного с потребностями дифференциации. А это означает, что здесь всегда будет компонента оксидативного стресса и радикалами, очевидно является всего лишь побочным проявлением от ограниченной возможности к самообновлению, восстановлению из-за низкого уровня митозов. Этот уровень старения относится к межклеточному уровню механизмов старения. На него накладывает свой отпечаток стадийное старение, которое способствует ему. Чисто клеточного, первичного уровня старения здесь нет, то есть того которое связано с эффектом Хейфлика.
В то же время в свете сказанного как можно дать объяснение по сохранению неотении у мужчины 32 лет, у которого признаки и болезни старости не проявлены?
Исходя из того, что старение может исходить из верхнего уровня стадийных регулировок, а также и из нижнего уровня программного старения клеток, следует выяснить при каком типе старения активируются и различные сигнальные пути, результатом которых становится высвобождение провоспалительных молекул? В патологический процесс вовлекаются все новые и новые клетки.
В свою очередь любое провоспаление по сути запускает те или иные линии обороны с итоговой целью: предотвращение оксидативного стресса. Но сенелит активируют и различные сигнальные пути, результатом которых становится высвобождение провоспалительных молекул. В патологический процесс вовлекаются все новые и новые клетки и предрасположенность к новым возрастным заболеваниям.
Митохондриальная недостаточность – важный элемент первичного старения на клеточном уровне.
Удобной моделью для анализа такого типа старения может быть поседение волос. Этот процесс некоторые авторы связывают с митохондриями, вырабатывающих энергию. Именно они как антенны наиболее чувствительны к возрастным перекосам. Известны случаи повторной пигментации волос. Это описано в случае пересадки печени или сшивания сосудистой системы старой мыши и молодой.
У стареющих клонов клеток в первую очередь страдают митохондрии. Но, тем не менее, это старение нельзя назвать исходящим первично из самих клеток. Ему способствуют внешние факторы. Следовательно, связывать его с эффектом Хейфлика = сугубо клеточным уровнем старения, не приходится.
Очевидно, старея, человек тем не менее не доживает до уровня, когда дальнейшим пределом ограничивающим его жизнь является утрата способности к делениям на клеточном уровне.
Таким образом, старение можно представить как двунаправленные потоки: с верхнего уровня связанного со стадийными изменениями = гормональным профилем и уровнем межклеточных взаимодействий. Сенелит нельзя приписать каким-то только генерализованным верхним процессам или сугубо на уровне клеток, или межклеточных взаимоотношений. Это взаимодействие верхних и нижних уровней, где частично процессу представлена автономия на клеточном уровне, но не меньшую роль играют стадийные изменения. Это и автономия процессов и централизация. Старение идёт и сверху, и снизу одновременно, причем все их моменты согласовываются, взаимодействуют, обуславливают.
Неотенический ювенил = неограниченная молодость.
Это когда организм по возрасту выходит за рамки ювенила, но при этом сохраняет все его признаки. Так черенки от ювенильных растений имеют намного большую потенцию к укоренению = регенерации корней. Это одно из свойств состояния ювенильности как всего маточного растения, так и всех его частей, то есть побегов. В мире животных неотенические формы тоже обладают высокой потенцией к регенерации.
Для большей доходчивости напомню, что в природе часто встречаются аномалии развития растений, например растение не переходит на проявление своих взрослых особенностей формирования кроны и побегов, а навсегда удерживает свои ювенильные признаки и во взрослом состоянии. Ярким примером тому могут быть декоративные формы хвойных, например у ели или у криптомерии. Такие формы сохраняют навсегда признаки ювенильности, что резко этим отличает их от нормальных растений. Тем самым они проявляют свою неотению.
Не лимитированный ювенил или возможность неограниченной ювенильности = неотении – универсальная реальность, присущая как миру растений, так и животных.
Действительно, подытоживая описания примеров неотении на растениях, аксолотлях и даже на человеке, можно сказать, что в каждом живом существе такая программа является универсальной и присуща всем. В мире живого это может происходить без перехода на программы зрелости и продвижения по стадиям онтогенеза как матура, сенелит. Стадийно неотенические формы ели проявляют не лимитированную = вечную молодость. Это же мы наблюдали и на примере аксолотля, голого землекопа и даже человека.
Для эволюции нет проблем создавать неотенический ювенил у различных видов. Но это остановит филогенез. Приспосабливаемость такого типа жизни будет резко ниже чем у генеративно размножающихся.
В тоже время легко может сочетаться неотения и возможность размножения генеративно. Это не мешает филогенезу. Во многих случаях природа пользуется этой возможностью, в том числе это особо выражено в популяции человека. В этом случае неотения сочетается с половозрелостью. Амблистомы аксолотлей могут пожизненно не трансформироваться в саламандру.
Неотенические формы среди растений.
Если присмотреться на фото выше к строению побегов у клонового дерева ювенильной формы ели, то можно увидеть, что побеги её по форме и дорсовентральному охвоению все схожи с 1-2-летними сеянцами ели, которые имеют только в этом возрасте особую ювенильную внешность, отличающую их от взрослых растений.
Таким образом, здесь мы видим типичный неотенический вид растения без ограничений по периодам и другим условиям сохранения ювенильности. Это так называемая «открытая програмно ювенильность». Онтостадийного старения здесь не бывает. Растение если и умирает, то только молодым.
Следовательно, старость, а точнее онтостадийный сенелит не обязательны, не неминуемы. Отмирают такие ювенильные растения только по причине внешних факторов, но никак не связано с сенелитом, матурой и адальтусом. Таким образом, прохождение через все стадийные этапы онтогенеза не неизбежно в природе, абсолютно возможно полное сохранение безграничной ювенильности.
При этом следует отличать понятия сенелит связанный с онтостадийностью и записанный програмно как стадия онтогенеза, от обычного старения связанного с внешними факторами и не исходящий изнутри.
Причем, в данном случае неограниченная ювенильность у ели связана явно с эпигенетичскими перестройками, а не с сугубо мутационными процессами. Это подтверждает пример на фото выше, когда из ювенильного экземпляра ели израстает стадийно взрослый побег. Значит генопрограмма на трансформацию = метаморфоз с ювенильной формы во взрослую, в зрелое растение не сломана, а заблокирована, возможно снижением порога чувствительности гормонального профиля к трансформации.
Важно, что состояние ювенильности является необходимым условием для блокирования = отключения программ зрелости или готовности к половому размножению. Ювенальность поддерживается активными механизмами, задерживающими наступление взрослого периода жизни.
Таким образом, на каждую стадию онтогенеза предусмотрена своя стадийная программа, которой если не мешать, то она будет работать неограниченно. Таким примером не выключенной программы ювенильности являются клоны ели с неограниченным морфогенезом ювенильности. Неотения у них может продолжаться десятилетия и изнутри на уровне программ этому нет помех. Только внешние факторы могут препятствовать жизненности этих растений.
Это в свою очередь показывает, что старение, а точнее онтостадийный сенелит не является неизбежным, неминуемым процессом. Растения могут неограниченно существовать и вне этих возрастных программ. Принципиально нет препятствий возможности существования неограниченно на ювенильной стадии, что означает что потенциально и реально неограниченная молодость возможна.
Онтогенез состоит из ступенчатой последовательности чередования ряда программ. Каждая программа может работать по безлимитному принципу. Но в природе предусмотрено так что каждая предшествующая программа закладывает в себе условия для подключения следующей.
Существуют программы, сугубо отвечающие за специфику морфогенеза на каждом этапе онтогенеза.
Но над всеми этими стадийными морфогенетическими программами существуют надпрограммы, а точнее квазипрограммы, которые переключают переход с одного этапа на другой. Существует два противоположно направленных процесса: старение и омоложение.
Неотения противоположное крыло процесса прогерии.
Прогерия у животных — это когда при остановке соматического роста = вегетирования у растений продолжает нарастать половое созревание и перезревание, или появляться иные признаки преждевременного старения, которое идёт во вред особи и приводит к её гибели. Наглядным примером этого является прогерия у рыбы горбуша, у которой при нересте в возрасте 9 лет происходят резкие патологические вредные для особи изменения гормонального профиля, что приводит к резкому её старению и неспособности питаться, а затем к массовой гибели всей популяции среди данного возраста. Такая мортальность = смертность запрограммирована природой, так как ей это целесообразно. Сделано это в целях филогенетики популяции. Здесь филогенез определяет = управляет = подбирает выбор того или иного типа сенелита = прогерии, который в данном случае является ускоренным, в жестких рамках программ.
Таким образом, эволюция в качестве одного из своих инструментов выбора для оптимизации жизнеспособности = живучести вида может воспользоваться в процессе филогенеза как неотенией = растянутой юности, так и прогерией – ускоренным жестким старением. Также филогенез может воспользоваться растянутым адальтусом, что особо выражено у человека. Зрелый и трудоспособный возраст у человека может растянуться от 20 до 70 лет, то есть около 50 лет. Фазы матуры и сенелита могут растянуться на 15 – 25 лет. Причем это возможно только у человека, как социального существа, которое имеет возможность сохранять старых особей.
Если расширить смысловую нагрузку термина прогерия, то его можно сравнить с процессом патологической матуры - перезревания у деревьев, когда полностью замедляется не только рост, но и самовосстановление побегов. Это может сопровождаться избыточным = непомерным заложением стробил – генеративных половых структур. Если адальтус это нормальная функция зрелости, то матура это её крайность, чрезмерность, идущая особи во вред.
Прогерия это более жесткое и преждевременное старение с выраженными чертами патологии, то есть патологический тип старения, тогда как матура это более устоявшийся процесс присущий последним этапам онтогенеза. Но суть, конечные результаты и механизмы их имеют принципиальное сходство.
У растений можно признать как прогерию появление среди многолетних видов их же однолетних подвидов. Однолетние травы с циклом воспроизводства семян в первый же год часто имеют ряд преимуществ по выживаемости на пастбищах, где выгоняется скот.
Отличие сенелита от матуры.
Оно проявляется в том, что процессы самовосстановления переходят за планку уравновешивания и становятся слабее = отстают от процессов самоотмирания. Это истощает последние жизненные силы дерева, резко тормозит их вегетативную фазу, ослабляет Витаукт и ведёт к сенелиту.
У человека перезревание = матура проявляется в чрезмерности признаков зрелости = адальтуса. Так происходит климакс, а также чрезмерный гирсутизм = отрастание огромной бороды и оволоснения, аналогично тому, как это происходит у рыб горбуша, у которых в фазе матуры изменяется = разрастается челюсть = изменение её прикуса = прогения. Все эти признаки функционально не только бессмысленны, но и мешают жизни.
Старость — это болезнь или болезни старости - это что?
Избыточный провоспалительный или гормональный перекос являются плацдармом для проявления множества возрастных болезней = проблем. Как правильно их трактовать – болезни старости или просто болезни существует самый различный спектр мнений. Но следует признать, что существует патологическая старость, генетически обусловленная, а нормальное старение = матура тоже предрасполагают и несут в себе корни болезней.
Существуют ли универсальные закономерности ювенила, адальтуса, матуры и старения?
Анализ всех возможных случаев онтогенетического прохождения стадий развития показывает, что таких строгих универсальных принципов нет.
Для каждого биологического вида существует своя модель развития, в том числе для сенелита и мортуса = отмирания.
НЕОТЕНИЯ = ПРОДЛЕННЫЙ ЮВЕНИЛ В МИРЕ НАСЕКОМЫХ.
У насекомых важной частью их онтогенеза является метаморфоз, когда личиночные стадии развития или гусеницы резко морфологически отличаются от взрослых форм, у которых могут появится крылья и конечности.
Интерес представляют маточные личинки. Они проявляют не полный метаморфоз и продолжают сохранять некоторые ювенильные признаки = ювенильную неотению во взрослом состоянии. Это позволяет взрослым особям продлевать длительность жизни многократно, например у пчел в десятки раз.
Обычные особи при метаморфозе могут из гусениц перейти в существо с крыльями и конечностями, и иметь ограниченный рост. У маток многое по-другому. Они проявляют высокую половозрелость в плане плодовитости, но сохраняют ряд признаков эмбриональности, в том числе и неограниченный рост и значительно большую продолжительность жизни. Своеобразная неотения, но у насекомых. Причем генеративное перезревание не мешает им одновременно сохранять и элементы эмбриональности.
Наиболее выраженно это происходит у термитов. У термитов выражена кастовая принадлежность. Причем она в меньшей степени связана с особенностью питания, а предопределена генетически. Судьба насекомого предопределена уже со второй линьки, когда становится понятно по какому пути оно пойдет: по пути нимфы или рабочего-солдата. Причем переход с одного пути на другой уже жестко закрыт. У нимф образуются крылья и со временем они способны улететь для образования новой колонии. Можно сказать точнее, что она не предопределена, а закреплена как часть онтогенетической программы, стадия, наподобие циклофизиса у растений, проявления которого жестко связаны и с клоновыми поколениями.
В случае гибели размножающейся царицы или царя их роль «и.о. царя и царицы» могут взять на себя как рабочие бескрылые самцы и самки, так и нимфы с зачатками крыльев. Они приступают к размножению, сохраняя внешний вид ювенильных (неполовозрелых) особей. Таким образом неотения здесь предусмотрена природой. Длительность жизни рабочих и солдат 1-2 года, тогда как неотенические формы живут 30 лет.
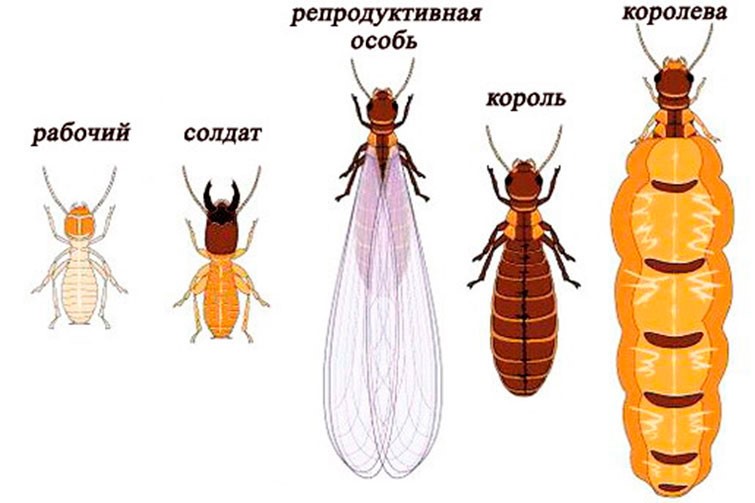
На рисунке: морфологические разнообразие особей среди термитов, которое существенно влияет на длительность онтогенеза.
Таким образом, у насекомых, в отличие от мира растений, генеративная перезрелость не закрывает вегетативные механизмы роста и поэтому не является началом адальтуса и сенелита. Следовательно генеративная зрелость не является обязательным механизмом, перекрывающим вегетативную мощь.
В свою очередь пример с галапагосскими черепахами показывает, что они могут жить очень долго, около 200 лет, но при этом проявлять очень медленный рост. Следовательно темпы роста в мире животных необязательный признак ювенильности. Здесь правят другие правила, которые связаны со способностью поддерживать длительно активность стволовых клеток.
НЕОТЕНИЯ = ПРОДЛЕННЫЙ ЮВЕНИЛ В МИРЕ ЖИВОТНЫХ.
Голый землекоп – лучший пример из мира животных проявляющий неотению.

На фото: Голый землекоп – рекордсмен по продолжительности жизни в своей весовой категории, может жить до 30 лет.
Животное, которое живет в 10 раз больше, чем мышь из той же весовой категории. Связано это с генетической программой на долгожительство и обеспечивается это механизмами неотении.
Именно эти механизмы позволяют ему противостоять болезням старости:
- онкологическим;
- сердечно-сосудистым;
- нейродегенеративным.
Дают ему эти способности не болеть высокая способности к самоомоложению. Используя силы неотении, присущие голому землекопу, человек мог бы жить 1000 лет.
Особенностью землекопов является чрезвычайно длительный период созревания мозга: их мозг долго остаётся в незрелом ювенильном состоянии.
Почему это важно для продолжительности жизни? Уточню что у человека старение начинается с мозга, именно он задаёт начало и темпы старения. Вернее, стареет столько не сам мозг, а его гормонально-вегетативная часть. Именно мозг является дирижером темпов старения. Если бы мозг так быстро не изменялся, то продолжительность жизни человека можно было бы увеличить значительно.
В первую очередь тон задают и определяют не работа всех стволовых клеток организма, а именно состояние мозга. Ситуация здесь аналогична тому, что и в мире растений, где мы уже выше показали, что у деревьев в кроне имеется разнокачественность по темпам старения. Стареют в первую очередь плагиотрпные побеги, в которых наблюдается полный перекос в гормональном профиле, а сенелит у побегов из апикальных и спящих почек может не проявляться вообще, там он не обязателен. Тем не менее, он может проявляться в программах у короткоживущих видов растений. Природой филогенетически предусмотрена возможность существования видов короткоживущих – это необходимо там, где высокая степень конкуренции среди множества видов и преимущества по сохранению вида могут получить короткоживущие виды.
Землекопы явно относятся к той группе существ, которым природой генетически предусмотрено долгожительство: с возрастом они не дряхлеют, не лысеют, не теряют зрение и зубы, да и не знают старческих болячек. Неудивительно, так как живут они в такой экологической нише (под землёй), где нет конкурентов.
В чем проявляется неотения землекопа? Если просмотреть картинку, где изображены этапы развития крысы, то легко можно отметить, что новорожденный крысёныш имеет полное сходство и отсутствие оволоснения как и у взрослого землекопа. Признаки ювенила сохранены и у взрослой особи.

На рисунке: у взрослой особи землекопа сохранены бесшёрстные признаки ювенила = неотении, как и у новорожденной крысы.
Ниже предложено фото обезьяны в детском виде и во взрослом состоянии, из которого становится очевидным куда идёт эволюция человека – в сторону сохранения всех лучших черт неотении.

На фото: слева неотенический облик детёныша шимпанзе, близкий к человеческим чертам, а справа взрослая особь. Из чего становится очевидным, что эволюция человека шла по пути неотении или педоморфоза.
Если сравнить длительность жизни человека с обезьяной, то можно тоже отметить эту же закономерность – резкое увеличение длительности жизни у человека. Если мы посмотрим фото детёныша обезьяны с человекообразным обликом и её взрослой особи, то легко убедиться, что эволюция человека пошла по пути сохранения лучших черт неотении.
Возможна ли полная реювенилизация организма путём включения механизмов неотении?
Особенности неотении на примере аксолотлей.
Выраженное явление неотении можно наблюдать у аксолотлей. Это — неотенические личинки амбистом — одного из семейств саламандр. Здесь в этой группе животных неотения — это специфическая особенность организма, при которой детёныши имеют полностью развитые половые органы и им необязательно приобретать признаки взрослой особи для продолжения рода. Это своеобразное сохранение детскости без последующих этапов онтогенеза. При этом личиночная фаза амблистом аналогична личиночной фазе у насекомых. Неотения связана с недоразвитием щитовидной железы. Она практически не синтезирует йод, который выполняет функцию активатора метаморфоза.
Но при этом их особенность в том, что они, будучи взрослыми способными к размножению, тем не менее обладают эмбрионоподобными признаками. Причем они обладают невероятно высокой степенью к регенерации, вплоть до восстановления конечности, кожного покрова, некоторые внутренние органы, позвоночник и даже участки мозга!
Аксолотли обладают достаточно высокой продолжительностью жизни – до 15 лет. Аксолотли могут превращаться в амбистомы и во взрослую особь в условиях изменения среды в виде уменьшении влажности, пересыхания водоёма. В этом случае они утрачивают наружные жабры и переходят на обычное сухопутное дыхание. Также на это может повлиять простое добавление в пищу гормона щитовидной железы - тиреоидина. Так их можно "метаморфизировать" - превратить в амбистом. Пара месяцев замачивания мотыля перед кормлением в растворе тиреоидина позволят, чтобы аксолотли из водных существ приняли взрослую форму и далее жили на суше как саламандры.
В тоже время есть данные, что метаморфоз у некоторых видов можно достичь обратным действием, то есть через снижение уровня выработки гормона тиреотропина. Для этого обычно нужны условия с нехваткой кислорода и пониженными пищевыми ресурсами адаптивно которым подходит низкий уровень гормона. Для форм активных нужен повышенный уровень. Все эти требования адаптации закрепляются у разных видов генетически. Но если генетических закреплений нет, то морфогенез можно достичь искусственно, например добавляя его в пищу или среду обитания или же уменьшая его содержание.

На фото: Аксолотль, ювенильная неотеническая форма.

На фото: Взрослая форма саламандры.
Каковы возможности реювенилизации с помощью питания на основе продуктов из меристем в сочетании с голоданиями?
Выше мы обсуждали возможные методы реювенилизации с помощью питания на основе продуктов с меристемами, имеющих сверхвысокий инициирующий биопотенциал. Для усиления результата его проводят с чередующимся голоданием с целью аутофагии клеток мешающих росту более здоровых линий, а также сенолитики, то есть устранением аутолизом (саморассасыванием) стареющих линий клеток. Именно эти сенелирующие линии превалирует над процессами омоложения и репарации.
Это подобно тому, что мы разбирали на примере хвойного дерева, когда накопление чрезмерного количества плагиотропных побегов тормозит рост апикальных центральных побегов и ведет к включению программ на сенелит. Есть приёмы омолаживания деревьев обрезками. В данном случае голодание действует аналогично обрезкам.
Особенности высших животных в том, что их регулировка онтогенеза завязана на уровне эпифиз-гипоталамус.
Тем не менее, радикально изменить ситуацию приведенные методы по употреблению меристемной пищи и голоданию не позволит, но обеспечит определенные оздоровительные моменты. Таким образом, всё это работает поверхностно в пределах возможностей коридора гормезиса, но не отменит генетические коды, блоки. Надо идти глубже, уметь обходить пределы, обеспеченные генетическими программами, например, связанные с переходом на новые этапы онтогенеза, последняя из которых запускает сенелит.
Продление молодости: задача не допустить становления последних этапов онтогенеза путём растягивания состояния неотении.
Так доказано статистически, что чем позже наступает фаза климакса, то есть возрастных гормональных перестроек, тем дольше жизнь.
Это позволит сохранить состояние ювенильности у взрослого человека, а значит повышенной способности к регенерации органов, отмене чрезмерного гормонального перезревания, отмене ряда симптомов старения как седина, дряблость, возрастные болезни к которым относятся атеросклероз, слабая память, снижение работоспособности.
Сложность высших животных организмов в их глубокой блокировке обратных переходов на этап ювенила.
Среди высших животных переходы на обратные этапы онтогенеза не могут регулироваться внешними факторами. Их значимость сводится к нулю. Переходы закреплены в филогенезе генетически. Задача идти дальше, вглубь, научиться управлять и переключать генетические программы-ограничители.
Следует учитывать, что переключение генетических программ на неотению (= ювенил) и обратно на имаго (= адальтус, зрелость) всегда связано с первичными переключениями в высших нейрогормональных системах регулировок. По сути, они заменяют роль апекса у растений. У некоторых низших животных (земноводные) регулировка зрелости осуществляется через уровень гормона щитовидной железы. У высших видов оно опосредуется через работу гипоталамуса и гипофиза, где запускаются или тормозятся механизмы не только выработки тиреотропного гормона, влияющего на работу щитовидной железы, но и многих других. Это подсказывает, что для реювенилизации надо искать механизмы, действующие через гипоталамус. В свою очередь это указывает на необходимость действия через эпифиз и все соподчинённые через него системы и оси регулировки. Эпифиз здесь дирижер, который завязан не только на циркадные циклы сна и бодрствования, но и на онтогенетические возрастные циклы становления организма.
Также как и в растении работа и уровень ответа главного апекса зависит от суммы альтернативно действующих сигналов со всех других точек роста, которые и определяют уровень старения апекса, так и в организме животного работу эпифиза определяет состояние развития периферии и желез-мишеней и соответственно гомеореза (гомеостаз в развитии) гормонального профиля. Гомеорез это гомеостаз гормонального профиля в динамике, развитии. Последний не может быть застывшим полностью, так как завязан на онтогенез.
Тут то и возникает масса вопросов и поставленных задач с целью разобраться с возможностями регулировки механизмов сна и технологий актиноритмов (соотношение продолжительности периодов освещенности и темноты) на эпифиз с целью его возврата в исходное состояние.
Все многообразие сигналов, обрабатываемых нервной системой, транслируется гипоталамусом в гормоны, в том числе и в тиреотропин-релизинг-гормон (ТРГ), стимулирующий гипофиз к синтезу тиреотропного гормона.
Способности к ювенильной регенерации у млекопитающих закрываются на этапе эмбриональной диапаузы.
Итак, высокие способности к регенерации проявляются у амфибий, например, у аксолотлей. Но эта же способность у высших животных, то есть млекопитающих, закрыта и, очевидно, на генетическом уровне она заблокирована уже на этапе эмбриональной диапаузы. Открыть эти гены можно только если залезть в зону этих программ. Как их разблокировать? Очевидно для этого нужны некие гормоны и факторы, которые имеются на фазе эмбриогенеза яиц или других эмбрионов. В последующие этапы эти гормоны исчезают, а программы, их обеспечивающие, наглухо глушатся. Это значит утрачивается способность к становлению тканей или их регенерации. Это позволяет предположить, что для этих целей можно использовать яйца птиц, но активированных в инкубаторах для возбуждения до ранних фаз эмбрионального развития, то есть на определенном промежутке диапаузы, когда еще вырабатываются морфогенетические факторы становления органов.
На фото: регенерация лапки у аксолотля

На фото: регенерация хвоста у ящерицы.
Возможность регенерации органов у людей.
Аксолотль — доказуемая модель по регенерации, способная воссоздать практически любую часть тела, включая мозг, глаза, сердце, центральную нервную систему. Это позволяет сделать вывод, что у людей есть «нереализуемая» возможность к регенерации.
Передача сигналов иммунных клеток — макрофагов — у аксолотля и мыши была сходной, когда они подвергались воздействию бактерий, грибов и вирусов. Ситуация изменилась, когда дело дошло до физического повреждения.
Оказалось, что макрофаги способствуют росту тканевых клеток у саламандры. Но в организме мышей макрофаги инициируют запуск процесса рубцевания, то есть только региональные механизмы. Более глобальные процессы были заблокированы. Предполагают, что шрам на месте травмы создаёт барьер для регенерации.
Предполагаю, что блокировка механизмов к рубцеванию позволит разблокировать более универсальные программы регенерации. В данном случае региональные программы ткани перекрывают более общие программы регенерации.
Неотеническая регенерация важная способность к реювенилизации.
Проблема в том, что с возрастом многие наши системы и органы выходят из строя. В первую очередь мы теряем большинство своих зубов, страдают и вырождаются ткани почек, где вместо почечной ткани появляются заместительные, соединительнотканные, своеобразный нефроз почек. Гибнут и не восстанавливаются нейроны, по некоторым данным ежедневно их гибнет до десяти тысяч. Страдают клетки поджелудочной железы, что ведёт к диабету. Дистрофия сетчатки глаза тоже связана с гибелью клеток, те же процессы идут в печени, эндотелии сосудов, эпителии кожи и т.д. Ни одна из этих тканей затем не может восстановиться полноценно никогда. Возможности к регенерации у высших животных закрыты уже на этапе эмбриональной диапаузы. Обеспечивают становление этих тканей в эмбриогенезе специальные тотипотентные белки, которые и открывают гены морфогенеза. Позже эти тотипотентные белки (работают на начальных этапах эмбриогенеза закладки тканей), а затем и фетальные белки исчезают (работают на последних этапах эмбриогенеза, то есть, где преобладают рост и дальнейшая дифференциация). Очевидно их можно все же запустить с помощью ряда приёмов, в том числе и с помощью живых пробужденных яиц, которые и принимают в виде коктейля.
У высших позвоночных животных регенерация органов практически полностью закрыта, но тем не менее есть и исключения из правил.
Итак, эти тотипотентные белки и стволовые клетки в принципе можно запустить и у взрослых зрелых особей, если открыть соответствующие генетические программы. В природе они находятся под контролем программ онтогенеза и закрыты, но тем не менее если они управляются биохимическими путями через тотипотентные белки, то есть основания быть уверенным, что этого же с их помощью можно достичь и у человека.
У саламандр после ампутации конечности рана быстро затягивается эпидермисом, а под ним формируется «шапочка» из недифференцированных клеток — бластема. Откуда берутся эти клетки? Известно, что многие клетки в районе ампутации гибнут, а оставшиеся дедифференцируются. Например, многоядерные клетки скелетных мышц распадаются на одноядерные клетки, а потомки этих одноядерных клеток, возможно, могут превращаться в фибробласты — клетки соединительной ткани. Но насколько они плюрипотентны? В костном мозге, мышцах и соединительной ткани есть и недифференцированные, стволовые клетки. Но насколько важен их вклад в регенерацию?
Проявление генетически врожденной или сохранение приобретенной = не наследственной неотении возможно как у растений, насекомых, так и у животных.
Это подтверждает проведенный нами анализ изложенного по самым различным формам жизни. Неотению можно поддерживать практически в любой форме проявления жизни. Очевидно, отмычкой к разгадке тайны природы будет понятие сохранение вегетативной силы.
Если мы будем плохо питать организм человека, то он, как и на примере дерева лиственницы тоже будет ускоренно созревать, а значит и ускорять наступление этапа старения. Если мы будем усиленно питать этот организм, то приведем к ускоренному росту или ожирению и незначительной задержке репродуктивной фазы и всей её симптоматики. Задача чтобы вегетативная фаза преобладала над генеративной, но и её возможности были ограничены, уравновешены возвратными процессами.
Это возможно осуществить на фоне резкого ограничения приёма углеводов, насыщенных жиров и некоторого уменьшения белков. Но главное всё это осуществляется на фоне приёма большого количества биогенных стимуляторов как пчелиное маточное молочко, плацентарные продукты, зародыши растений, яиц и им подобных.
Напомню, что пчелиная матка питается не медом, как это происходит у рабочих личинок, то есть энергетическим субстратом, а исключительно маточным молочком, которое в ней на долго сохраняет состояние неотении и продлевает тем самым её длительность жизни в десятки раз. При этом репродуктивная способность у неё не снижается. Тоже самое наблюдается и у термитов, размеры маток которых увеличиваются по сравнению с рабочими особями в 10-ки раз.
У человека такого увеличения роста и размеров организма не происходит, но это тем не менее заменяется на восстановление гормонального профиля, нейромедиаторного фона, в частности усиления работы эпифиза и восстановления выделения им мелатонина, как и в ювенильной фазе, то есть в молодости и детстве. Такой гормональный профиль удерживает на высоком уровне Жизненную Силу организма, его регенерационные и репарационные возможности, и способен существенно продвигать продолжительность его жизни так же, как и у маток термитов.
В отличие от насекомых, в человеческом организме рост ограничивается созреванием эпифиза костей, то есть их ростовых зачатков.
НЕОТЕНИЯ И ПРОГЕРИЯ У РЫБ.
То, что неотения возможна у рыб нет никаких сомнений, так как и у них и, например у аксолотлей она регулируется одинаково, то есть уровнем гормона щитовидной железы. Особенностью многих рыб является открытый тип роста, когда с возрастом происходит равномерное линейное увеличение пропорций и никаких качественных изменений в морфогенезе. Как внешняя форма существа, так и степень окостенения хряща и другие параметры сохраняются и у молоди, и у взрослых.
Кроме этого, у них имеется и прогерия, когда при остановке соматического роста продолжает нарастать половое созревание и перезревание, которое идет во вред особи и приводит к ее гибели. Это аналог процесса матуры - перезревания у деревьев, когда полностью замедляется рост, но происходит чрезмерное заложение стробил – генеративных половых структур. Это истощает последние жизненные силы дерева, резко тормозит их вегетативную фазу, ослабляет Витаукт и ведет к сенелиту.
Аналогом такой матуры является пример старения = сенелита и мортального исхода = смерти рыбы горбуша, которая живет от 1,5 до 9 лет и гибнет при единственном в её жизни нересте из-за гормонального перезревания и чрезмерной работы гипофиза.

На фото: Рыба горбуша, где нижний экземпляр находится на стадии крайней матуры и сенелита.
«ЭЛИКСИР МОЛОДОСТИ» - ВОЗМОЖЕН ЛИ?
Возможный механизм действия препаратов против старения.
Чем старше становится человек, тем больше в тканях его организма накапливается стареющих клеток. Это, похоже, способствует развитию ряда болезней, например артрит и диабет. Если мы научимся разрушать эти клетки или избавляться от них, то это позволит повернуть процесс старения вспять. В состав разрабатываемого вещества входит пептид, который побуждает стареющие клетки к самоуничтожению.
У старых мышей после применения препарата возросла физическая выносливость, животные стали больше времени проводить, бегая в «колесе», а расстояние, которое животные преодолевали во время испытаний, после приема «эликсира молодости» выросло практически вдвое.
Выявлено улучшение работы внутренних органов мышей. Даже шерсть подопытных грызунов стала выглядеть лучше.
Предполагают, что созданный препарат может быть эффективен в борьбе с раком.
Фактор молодости из плазмы.
Очевидно, плазма молодых доноров содержит большое количество молодых клеток и присутствуют стволовые клетки, которые способны выделять факторы способные восстанавливать угасающие функции.
Найдено, что стимулятором регенерации мышечных клеток может служить «гормон любви и привязанности» окситоцин. Очевидно, что каждый тип ткани имеет свои ключи = белки, определяющие их молодость. Так например, за восстановление нервных клеток отвечает активность гена TET2. Мутации в этом гене могут повлечь за собой не только когнитивный упадок, но и другие «возрастные» заболевания, включая рак, сердечно-сосудистые нарушения и инсульт.
Похоже «разлад» в гене TET2 можно устранить, разбавив старую кровь молодой, богатой особыми регуляторными ферментами. Существует много кандидатов на роль «главного фактора молодости».
Метод ифузии = переливание крови от молодых к старым и её недостатки.
Главный недостаток – кратковременность действия. Единичные вливания белков и других компонентов молодой крови вряд ли могут изменить функционирование пожилого организма. Похоже, что, попав в новую среду, эти чудо-протеины заставляют тело пациента вырабатывать их самостоятельно. Но весомых доказательств этой гипотезы пока нет.
Также известно, что в старой крови циркулируют некие вещества, которые действуют противоположно ювенилизирующим. Это вещества блокторы регенерационных процессов. Следовательно старость это не просто недостаток некоторых омолаживающих веществ, но и одновременно присутствие специфических веществ старения.
В организме человека разного возраста находится разное соотношение омолаживающих и старящих веществ. Всё зависит от их соотношения.
Поэтому для закрепления эффекта омоложения процедуру переливания молодой крови потребуется регулярно повторять.
Существуют противоречивые мнения по поводу инфузий крови: побочных явлений нет, но другие допускают возможность аллергических и других аутоиммунных проявлений.
Допускается возможность того, что активация стволовых клеток, вызванная вливанием молодой крови, окажется канцерогенной. Возможно в плазме людей-доноров могут случайно оказаться не только «белки молодости», но и неблагоприятные факторы, способствующие развитию мышечной дистрофии, болезни Альцгеймера и других недугов.
Поэтому, пока эта процедура представляет собой неоправданные риски при отсутствии достоверных эффектов. Механизм влияния трансфузии молодой крови на увеличение продолжительности жизни пожилого человека и улучшение его мозговой деятельности до сих пор не понятен.
ЭВОЛЮЦИЯ ВНЕШНИХ ЧЕРТ ЧЕЛОВЕКА ИДЁТ ПО ПУТИ НЕОТЕНИИ.
Так в эксперименте если предложить молодым девушкам колоду из фото людей различного возраста и внешнего вида, как на фото: тип 1 и тип 2, то привлекательнее в большинстве случаев будут типы с более мягкими чертами внешности, а не с грубыми чертами, сильным оволоснением и агрессивной психологией.

На фото: представлены два мужчины одного возраста. У мужчины слева отсутствуют вторичные пубертатные признаки – борода и усы. У мужчины справа - обычный тип пубертата.
Среди азиатов эволюция продвинута дальше, так как среди них особо часто встречаются безбородые мужчины. Неотения распространена среди азиатов благодаря мутации гена EDAR.

На рисунке: Облик молодой влюбленной пары с умиляющими всех чертами молодости и нежности чувств. Вот истинный смысл жизни и движущая сила эволюции людей в сторону неотении.

На рисунке: Образы обаятельности и нежности в юности, которым отдадут своё предпочтение большинство девушек мечтающих создать семью, что содействует отбору неотенических линий.
Всмотритесь на рисунки людей, которые у большинства из нас вызывают чувства нежности, умиления, а у девочек симпатии. Таким обликам мы всегда отдаем предпочтение.

На рисунке: Современный эталон красоты облика мальчика, привлекающего свой обаятельностью. Любой идеальный эталон совпадает с чертами ювенильности.

На рисунке: Образы обаятельности и нежности в юности, которым отдадут своё предпочтение большинство девушек мечтающих создать семью, что содействует отбору неотенических линий.

На рисунке: Облики артистов молодых людей, кумиров, от которых в восторге большинство современных девушек. Лидируют только неотенические формы лица. Эволюционный отбор у людей идет по пути неотении.
ПРИМЕРЫ ЛЮДЕЙ, ДОСТИГШИХ РЕАЛЬНОГО ПРИОСТАНОВЛЕНИЯ СТАРЕНИЯ.
На фото показана женщина в возрасте 51 года, которой по внешности обычно дают 18 лет. По профессии библиотекарь, но активно занимающаяся своим телом с помощью активного образа жизни, плавания, увлекается моржеванием в холодной воде. Утверждает, что молодость её кожи связана с перепадом температур при водных процедурах.
На лице у этой женщины нет ни одной морщины, а ровный, красивый цвет ее лица говорит о том, что у нее нет проблем с кожей. Лю почти не пользуется косметикой, так как уверена, что в декоративных средствах содержится много вредных веществ, которые негативно сказываются на коже.

На фото: китаянка Лю в возрасте 51 года выглядит как 18-летняя.

На фото: Нежная кожа лица китаянки Лю в возрасте 51 года. Её возраст большинство определяет как 18 лет.
Она не ест мясо, употребляя овощи, зелень и фрукты. Из напитков она предпочитает зелёный чай и воду.
На фото показан мужчина 68 лет, которому почти никто не даёт более 30 лет.
На фото приведен облик Ху Хая 68-летнего жителя Шанхая. Увлекается физическими упражнениями, йогой, ЗОЖ.
Согласно медицинским анализам, большинство показателей (лейкоциты, хрупкость костей, состояние сосудов) соответствуют примерно 40-50 годам, что на 15-20 лет меньше его настоящего возраста.

На фото: приведен облик Ху Хая 68-летнего жителя Шанхая. Увлекается физическими упражнениями, йогой, ЗОЖ.

На фото: первое фото снято в 1978 году, а второе - в 2016, разница практически не заметна.

На фото: Поль Брэгг – известный популяризатор темы здоровья, автор книг, в том числе книги бестселлера: «Чудо голодания».

На фото: Поль Брэгг – в возрасте 82 года он выглядел как 40-летний.
СЛОВАРЬ ТЕРМИНОВ.
Адальтус – Этап онтогенеза. Имеет начальную полезную для организма функцию зрелости = полный пубертат, это, по сути, более повзрослевший этап развития, у людей это около 25-50 лет.
епатопения – уменьшение функциональных клеток в печени.
Фетальный – Этап онтогенеза. У растений молодые ростки, а у людей грудничковый период.
Имаго = постнатальный – Этап онтогенеза. Полностью развернутые программы морфогенетического становления организма, например у людей это соответствует подростковому периоду, но не с полным проявлением половых признаков, например отсутствие бороды = неполный пубертат.
Онтофизис – термин предложен Гарбузовым Г. А. – подразумевает не простое сохранение у клонов неких особенностей роста и плодоношения, а проявление более общего процесса, то есть сохранения степени онтостадийной зрелости. При этом расширено понимание этого явления не только для клонов, но и в пределах данной особи растения, когда новое поколение побегов в пределах одного и того же дерева все проявляют продвинутую физиологическую или морфологическую стадийную зрелость. Программы на ювенильность у них заблокированы.
Матура – Этап онтогенеза. Стадия перезрелости организма, например у растений это переизбыток закладки генеративных почек, резкое или полное торможение вегетативной мощи. Таким образом это генетическая программа на чрезмерность, идущая особи во вред; а у людей это совпадает с постклимактерическим периодом, связанным с резким изменением гормонального профиля
Сенелит – Этап онтогенеза. Стадия появление как первых признаков старения, обозначаемых как патологическая врожденная синдроматика старения.
Сенесцент – Старение на нижних этажах клеток и тканей.
Саркопения – мышечное вырождение, атрофия.
Меланопения - седые волосы = исчезновения стволовых клеток, вырабатывающих краситель меланин.
Нейропения – уменьшение нервных клеток.
Эпителиопения - эпителиальная дисфункция стенок сосудов, ведущая к атеросклерозу, отмирание до 50% микрокапилляров.
Топофизис - характер роста, то есть сохранение тех особенностей роста у клонов, которые они имели в кроне: ограниченный, слабый темп роста как у ветви.
Циклофизис – когда клоны могут сохранять те особенности кроны, которые связаны с проявлением его генеративной готовности = зрелости побега, то есть, например закладки стробил у хвойных или цветочных почек у лиственных.
Плагиотропный - горизонтальный рост, как растут ветви у хвойного дерева.
Ортотропный – способность роста побегов только вверх.
Ювенильный = натальный – Этап онтогенеза. Детство и юность без признаков пубертата = половой зрелости.

Похожие статьи
Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне