
Сиртуины - ключ к молодости на клеточном уровне
Что такое сиртуины (сиртуининг*) и противостоят ли они SASP (саспингу*)?
Сиртуины – семейство белков, обладающих деацетилазной активностью. Составляют класс гистоновых деацетилаз. Считают что они помогают защитить наш мозг и сосуды от старения, а также от рака. Есть мнение они способствуют активации Генов Молодости. Предполагается, что сиртуины регулируют процессы старения, транскрипции, апоптоза и сопротивляемость стрессу… Их связывают с увеличением продолжительности жизни.
Сиртуины* задерживают клеточное старение и продлевают жизнь организма через регуляцию различных клеточных процессов. Это происходит за счёт задержки возрастного укорочения теломер, поддержания целостности генома и стимулирования репарации ДНК.
Например, сиртуин 1 (Sirt1) предотвращает повреждение ДНК, остановку клеточного цикла, снижает окислительный стресс и старение эндотелиальных клеток артерий посредством ингибирования пути Forkhead box O. Таким образом сиртуины могут остановить клеточное старение.
Эффект продления жизни, оказываемый сиртуином-1, опосредован аутофагией.
Сиртуины – ключ к ювентальному реверсу*
Описанный ракурс по сиртуинам позволяет утверждать, что они могут иметь отношение к ювентальному реверсу. Реализуется за счёт сиртуининга* и противодействия саспингу*.
Исходя из обсуждаемой концепции способности сиртуинов к ювентализации клеток и противодействия сенесценту* Гарбузов Г.А. расширил понимание этой концепции и считает, что сиртуины можно рассматривать как аналог гормона цитокинина у растений. Это означает, что они обладают способностью возвращать стволовым клеткам плюрипотентный уровень. Но, тем не менее, они не позволяют достичь полной степени омоложения клеток, то есть достигнуть тотипотентной степени омоложения стволовых клеток.
Рассмотрим в чем аналогия сиртуинов с универсальным омолаживающим гормоном цитокинином у растений.
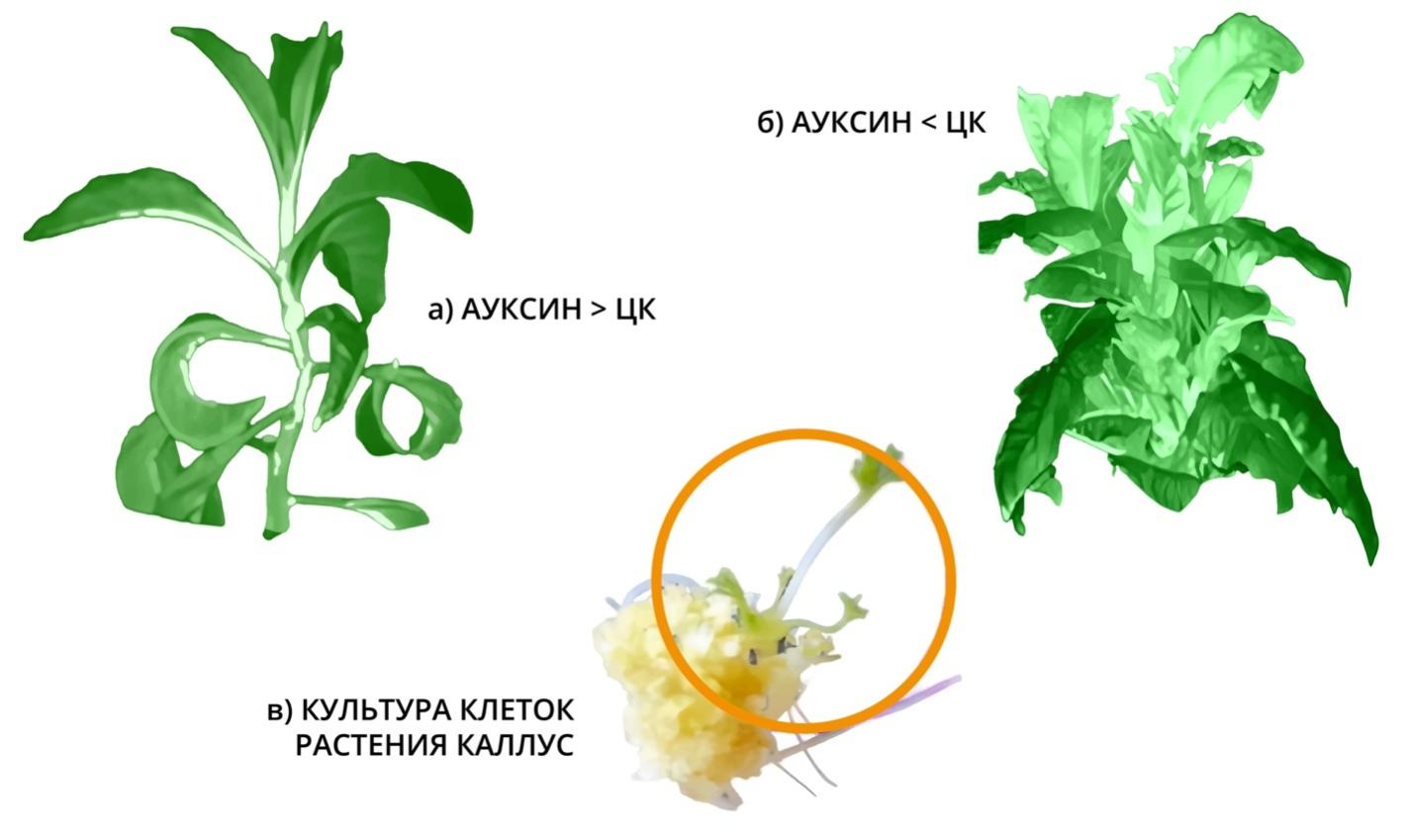 Рис. 1. На рисунке изображены результаты физиологического влияния растительного гормона молодости — цитокинина, где:
Рис. 1. На рисунке изображены результаты физиологического влияния растительного гормона молодости — цитокинина, где:
а) Растение без обработки ЦК.
б) Растение после обработки, которое поменяло свои морфологические признаки онтостадийной зрелости на более ювенильные формы.
Цитокинин – это гормон, который вырабатывается только у растений в их ростовых апексах корней. От уровня этого центрального гормона зависит всё благополучие растения, его Жизненная Сила, репродуктивная зрелость, способность к закладке не просто новых точек роста (почек), а даже к закладке эмбриональных зачатков. Внизу рис. 1. видно как из каллусной ткани клеток в культуре ткани при условии преобладающего действия цитокинина зарождается эмбриональный росток со всеми потенциями тотипотентности. То есть закладка не какой-то части организма, а именно заново всего организма. У человека такая тотипотентность исключена и замещена на плюрипотентность. Организм человека не может восстанавливаться тотипотентно, так как это бы противоречило его основным принципам существования – гомойотопию – ограниченности размеров и формы. Любое обновление и омоложение организма человека возможно только в условиях смены (устранения) одних линий клеток (поколений) и замены на их месте новыми. Для этого в нашем организме реализованы механизмы аутофагии и апоптоза. У нас есть точки «замены», точки самовосстановления, но не точки роста. Каждая ткань организма насыщена этими точками смены клеточных линий, здесь и осуществляется репарация ткани. Эти точки должны выделять некие гормоны для самовосстановления. Здесь это осуществляется с помощью сиртуинов. Это позволило автору выдвинуть концепцию, что они являются аналогами цитокинов, а значит через эти сиртуины можно и нужно манипулировать с целью усиления возможностей омоложения организма на уровне клеток и тканей. Чем больше вырабатывается сиртуинов, тем лучше и быстрее обновляются, омолаживаются ткани.
Итак, автор выдвигает концепцию, что сиртуины это аналог цитокининов. Но в организме растений цитокинины сами по себе не смогут вести регулировку по пролонгированному росту растения. Они обязательно взаимодействуют с ауксинами. Именно баланс их пропорций определяет зрелость или ювенильность, регенерацию и закладку новых почек. От степени их пропорций определяется возможность закладки самых различных типов почек: эмбриональных (закладываются вегетативные почки из которых разовьётся целое полноценное растение), спящих (из них образуются порослевые побеги обладающие воссоздать растение на прежнем корню со всем комплексом фенотипа ювенильного растения выросшего из семени), брахиальные, апикальные, репродуктивные и многие другие типы почек. Причем самыми зрелыми являются репродуктивные и брахиальные (пазушные) почки. Появление таких почек является результатом смены гормонального доминирования когда действие цитокининов прекращает проявлять свою доминирующую роль, а доминантность переходит к ауксиновым гормонам. Таким образом четко просматривается регулирующая ось: цитокины – ауксины.
В организме человека тоже очевидно имеется такая ось и на её роль автор выдвигает сиртуины – мелатонин. Но здесь имеется особенность, которой нет в растительном мире: у клеток животного типа предусмотрена коммитативно сенесцентность. Они имеют лимит репликаций или лимит Хейфлика = 52 делений. Это и обеспечивает неизбежную сенесцентность на клеточном уровне. Организм человека стареет с двух сторон-этажей: гормонального, где задающим дирижёром является эпифиз, и нижнего клеточного уровня с их лимитом Хейфлика. Скорость старения или величина онтодианной линии, реостат хроноса, или таймер на эпигенетических часах Стива Хорвата задаётся потенциями определёнными как с верхнего этажа (эпифиз), так и потенциями с нижнего клеточного этажа. Но движущей, реализующей силой здесь всё равно является ось Сиртуины ⇔ Мелатонин. Также как и у растений, где имеется выраженное взаимовлияние между цитокинином и ауксином, в организме человека имеется взаимокорректировка между сиртуином и мелатонином. Цитокинины у растения не могут в достаточной степени вырабатываться без соответствующего достаточного уровня ауксинов; а ауксины полностью зависят от мощности работы корневой системы и их цитокининов. В принципе многие растения показывают возможность неограниченного существования при соответствующем требовании к балансу этих гормонов. Секвойи или Сосна остистая могут существовать до 3000 – 5000 лет, а вегетативная роща Тополя осинолистного - до 85000 лет. Клоны большинства растений не имеют внутренних лимитов, то есть изначальной эндогенной коммитации (предписания) на ограниченное количество циклов репликаций. Работа их гена по активности фермента теломеразы, от которой зависит работа теломерного репликативного счётчика, не отключается. Поддерживают их непрерывную возобновляемость сиртуины. Но для этой высокой степени возобновляемости теломер нужна особая мощь и доминантная роль сиртуинов. Это возможно только в клетках имеющих потенцию пользоваться тотипотентностью, запускать тотипотентные программы. Следовательно, в клетках животного типа сиртуины (как гормоны) не имеют доминантного статуса. Высокую степень ювентализации клеток (снятия сенесцентного блока) сиртуины способны осуществлять только при особо высоких концентрациях! Каждая степень потентности клеток имеет различную степень уровня выработки сиртуинов. Каждая потентность имеет свою высоту порога сиртуинов, на который они становятся отзывчивыми или резистентными. Итак, не сами сиртуины определяют способность к ювентальному реверсу, а их уровень концентрации.
Очевидно эта ОСЬ и является рычагом потенциального регулирования в организме человека и позволяет подкручивать-корректировать наши биологические часы. Очевидно лимит Хейфлика основан на недостатке уровня выработки сиртуинов (ювентализирующий клеточный гормон), а в культуре ткани, то есть изолированных клетках, эти клетки теряют чувствительность, становятся резистентными к уровню мелатонина, который инициирует выработку сиртуинов. В данном случае можно утверждать, что сиртуины являются аналогами растительного гормона цитокинин, а мелатонин – аналогом гормона ауксин. Здесь в организме человека тоже работает принцип доминирования в оси, и наша ось сверху ослабевает из-за ослабления с возрастом выработки мелатонина практически в 10 раз. Это в свою очередь усиливает механизмы саспинга (процесс когда обычные и ювентальные клетки приобретают сенесцентный фенотип на основе сенесценс-ассоциированного секретома SASP), которые являются противоположностью механизмам сиртуинига. Итак, сиртуининг - путь к ювентальности, саспинг – к сенесценту. Также ось ослабевает и на нижнем клеточном уровне из-за отсутствия потенции на тотипотентность, которая бы позволяла развить более высокую мощь по выработке сиртуинов. В культурах клеток гены тотипотентности не могут быть запущены изначально, так как это исключено принципиально, а уровень плюрипотентности не позволяет обеспечивать достаточную мощь для выработки уровня сиртуинов. Принцип доминантности цитокининов = сиртуинов утрачивается, что не позволяет проявится как закладке полноценных апексного типа точек роста, так и ограничивает, ведёт к затуханию репликативных возможностей. Поэтому такая клеточная система приобретает ограничения на возможности полного самовосстановления. Эти ограничения идут как с верхнего этажа – эпифиза, так и с нижнего этажа коммитированный на детотипотентность (полная её блокировка).
И в тоже время сиртуины не несут в полной мере роль гормона, каковым является цитокинин. Особенность гормонов в том, что они вырабатываются в одной части организма и переносят сигнал на расстояние в другую часть. У растений цитокинин вырабатывается в корнях и по ксилеме поднимается в верхнюю часть растения и там обеспечивает свой эффект в виде перезакладки новых точек роста, обеспечивая растению наружный рост и изменение его формы. Организму животного такая роль гормона по перезакладке апексов не нужна, так как это противоречит его принципу существования, то есть сохранению размера и форм. Для животных важна не перезакладка точки роста, а её самообновление, для чего нужны другие гормоноподобные регуляторные вещества, которые работают в пределах каждой данной ткани организма. Такую роль здесь играют сиртуины. А гормонов, которые также обеспечивают возможность большой регенерации у животных нет. Поэтому основная ось самообновления тканей у животных строится не на ауксинах и цитокининах, а на других веществах: мелатонин – сиртуин.
Через цитокинин и его гипервысокие уровни у растений обеспечивается эффект так называемого радиксального доминирования* (когда превалирование ювенильного гормона корней определяет фенотипические особенности растения), которое обеспечивает растению состояние ювенильности и вегетации и может переводить в разряд долгожителей. В противовес цитокинину у растения стоит гормон ауксин, который вырабатывается только в условиях освещенности в центральных апексах. Мелатонин вырабатывается в темноте, но для его выработки из серотонина необходимо предварительное обязательное освещение.
Ауксин и мелатонин – функциональные прототипы. Через мелатонин в организме животных осуществляется эффект эпифизарного доминирования или эпифизарной молодости*, ювенильности. В период его доминирования подавляется репродуктивно-генеративная функция организма, но поддерживается вегетативно-ростовая компонента через гормон роста. Также ауксин - это индолуксусная кислота сходная с аминокислотой триптофан. Именно из триптофана строятся и серотонин, и мелатонин.
Кстати, у растений тоже имеются наравне с цитокининами и регулировка сиртуинами. Очевидно у растений регулировка сиртуинов осуществляется посредством цитокинина. У животных цитокинины исключены из этой системы и регулировка сохраняется только на месте, автономно через сиртуины. Функцию цитокининов частично замещает мелатонин. Но мелатонин не обладает потенцией к полной реювенилизации, как это возможно у цитокинина. У животных ювентализация возможна на уровне не ниже первой степени, то есть не допускается выход на тотипотентный уровень стволовости. Поэтому у них нет специализированного универсального для этого цитокининоподобного гормона. Однако у животных и растений сохранение ювенильности на уровне плюрипотентной потенции под силу уже справиться как ауксинам, так и мелатонину.
Через пролонгирование эффекта эпифизарной молодости появляется возможность повлиять на активирование работы сиртуиновой системы, повысить её ранг до сиртуинового доминирования, аналогичного цитокининовому радиксальному доминированию.
Что такое стем-сиртуиновое доминирование*?
Это когда преобладает стимуляция стволовыми клетками общего развития организма, то есть снизу, а не сверху. На последних этапах онтогенеза — матуры, адальтуса и сенилита, то есть в период инволюции гипоталамуса, его определяет принцип стем-соматического или прогениторного доминирования, когда исчерпаны возможности высокопотентных стволовых клеток, а доминируют соматические и концевые типы стволовых (прогениторные) с минимальной степенью потентности к репарациям, которые не позволяют происходить его дальнейшему самовосстановлению. Понятие «стем-доминирование» в биологии впервые ввёл Г.А. Гарбузов. Stem = стволовой.
Но стем-доминирование бывает как положительное, так и отрицательное и зависит от того на какой степени стволовости находятся стволовые клетки. Стем-доминирование сопряжено с активностью мелатонина или его неактивностью; в последнем случае ранг стволовости снижается и потенциал к ювентальному реверсу снижается. Стем-доминирование позволяет активировать и поддерживать выработку мелатонина в эпифизе. Угасание степени стволовости и стем доминирования приводит к переключению регулировок программ на онтодианной линии онтогенеза. В эмбриогенезе мелатонин не вырабатывается но стволовые клетки находятся в ранге плюрипотентности. В ювенильном возрасте уровень мелатонина максимальный и стволовые клетки имеют максимальную степень потенции, вплоть до 2 и 3-го порядков, в старости эта потентность резко снижается и не превышает уровень прогениторных = концевых стволовых клеток. Отсутствие мелатониновой поддержки на высоком уровне присущей юности и детству (но не эмбриональному периоду, когда не вырабатывается мелатонин) объясняет причину клеточного ограничения на репликации и самообновления, которое называют лимитом Хейфлика.
Очевидно «раскупорить» степень стволовости стволовых клеток можно меняя уровни мелатонина в организме до исходных в детстве до 12 лет. Это активизирует все точки самовосстановления и замены на новые клетки, и повысит тем самым уровень их функциональности и работоспособности. Это означает необходимость решения проблемы эпифизарной девиации — периода эпифизарного вечера, заката (отклонение от оптимального уровня активности, старости), который наступает после эпифизарного утра, эпифизарной молодости. (Термины предложил Г.А. Гарбузов).
Термины
- Саспинг – от слов SASPинг, то есть процесс когда обычные и ювентальные клетки приобретают сенесцентный фенотип на основе сенесценс-ассоциированного секретома SASP – термин предложен Г.А. Гарбузов.
- Сенесцент - репликативное старение, которое зарождается на уровне клеток.
- Сиртуины - белки-сиртуины являются одними из ключевых регуляторов клеточного метаболизма, функций митохондрий и ремоделирования хроматина.
- Сиртуининг – от слова сиртуины SIRTUIN – вещества содействующие омоложению, восстановлению клеток. Задерживают клеточное старение и продлевают жизнь организма через регуляцию различных клеточных процессов. Это происходит за счёт задержки возрастного укорочения теломер, поддержания целостности генома и стимулирования репарации ДНК – термин предложен Г.А. Гарбузов.
- Радиксальное доминирование - от слова «радикс» — корень, этап онтогенеза, когда доминирует гормон корней — цитокинин, который и определяет особенности этого онтогенетического этапа. Термин предложил Г.А. Гарбузов.
- Эпифизарная молодость - этап онтогенеза, который регулируется на принципах эпифизарного доминирования. Термин предложил Г.А. Гарбузов.
- Ювентальный реверс - возможность клеток в тканях возвращаться из зрелых и сенесцентных клеток в исходное стволовое состояние с преодолением лимита Хейфлика, а также проявлять обратный метаморфоз в ювенильное состояние – термин предложил Г.А. Гарбузов. Таким образом можно говорить о реверсе Гарбузова как противоположность лимиту Хейфлика.
 Книга "Эпифиз — верховный драйвер онтогенеза: ключ к молодости и старению"
Книга "Эпифиз — верховный драйвер онтогенеза: ключ к молодости и старению"
Геннадий Алексеевич Гарбузов — автор двадцати книг. В газетах и журналах Гарбузовым Г.А. опубликованы более ста статей на тему лечения хронических и трудноизлечимых болезней природными методами.
Старение — это многоуровневый процесс как на верхних этажах регулировок (системном), так и на нижнем клеточном. Чтобы адресно разрабатывать методы противодействия и сдерживания общего потока старения (геронтос), необходимо отыскать правильный ответ — откуда исходят истинные, первичные, начала этого общего процесса. Автором был проведён глубокий анализ природы старения как в растительном мире, так и в животном. Выявлено, что этот процесс является двусторонним: снизу он исходит на уровне клеток (сенесцент), причиной которого является эпигенетическая коммитация на основе онтофизиса, а также нисходит с верхних этажей, где верховным драйвером данного процесса является эпифиз, а не гипоталамус, как это считалось ранее. Опираясь на фундаментальные разработки, предложен комплекс методик воздействия на все уровни сдерживания процесса старения. Показана динамика взаимовлияния друг на друга верхнего и нижнего этажей старения. Именно верхний этаж определяет весь спектр возраст-зависимых заболеваний (анизомалий). Если мы научимся сохранять «эпифизарную молодость» = неотению, то отодвинем старость.
Книгу можно заказать на сайте: Garbuzov.org

Похожие статьи
Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне