
Эпифиз – верховный драйвер онтогенеза: ключ к молодости и старению
АННОТАЦИЯ:
Старение — это многоуровневый процесс как на верхних этажах регулировок (системном), так и на нижнем клеточном. Чтобы адресно разрабатывать методы противодействия и сдерживания общего потока старения (геронтос) необходимо найти правильный ответ – откуда исходят именно истинные первичные начала этого общего процесса. Автор провел глубокий анализ природы старения как в растительном мире, так и животном. Показано, что этот процесс является двусторонним: снизу он исходит с уровня клеток (сенесцент), причиной которого является эпигенетическая коммитация на основе онтофизиса, а также с верху, где верховным драйвером этого процесса является эпифиз, а не гипоталамус, как это считалось ранее. Исходя из этих фундаментальных разработок, предложен комплекс методик воздействия на все уровни сдерживания старения. Показана динамика взаимовлияния друг на друга верхнего и нижнего этажей старения. Именно верхний этаж определяет весь спектр возраст-зависимых заболеваний (анизомалий). Если мы научимся сохранять «эпифизарную молодость» = неотению, то отодвинем старость.
Книга I из 3-х томника: «БИОЛОГИЯ МОЛОДОСТИ»
Почему в гипоталамусе в первую очередь проявляются провоспалительные процессы как с болезнями, так с возрастом и старением?
Группа ученых во главе с Дуншэн Цай занималась поиском в мозговых структурах «центра старения». В ходе исследований специалисты в первую очередь обратили внимание на один из типичных симптомов старости — это завышенная чувствительность к разного рода воспалениям, что и стало критерием для дальнейших исследований. Ткани с провоспалением отличаются тем, что выделяют повышенное количество сигнального белка — так называемый фактор NF-kB, который включает клеточную реакцию на самовоспаление. Выявлено, что самая значительная концентрация белка NF-kB отмечалась именно в гипоталамусе — части самого мозга, управляющей деятельностью гормональной и вегетативной нервной системами. О чём это говорит? О том, что гипоталамус стареет быстрее всех и именно гипоталамус заводит темпы старения всего организма. Или другими словами его потенциал закрывается или ограничивается, или изнашивается быстрее других тканей. Это принципиально важный ответ, так как позволяет сделать заключение, что особенности различий по онтогенезу у различных видов животных, то есть длительности жизни, зависят от запаса прочности гипоталамуса, выступающего в роли камертона, настройщика темпов старения всех остальных тканей. Этот запас заложен эволюционно. Запас прочности обеспечивается генетическими механизмами противодействия провоспалению, а также особенностью возможностей длительного или неограниченного самовосстановления его стволовых клеток.
Уточню, что приведенные результаты эксперимента проводились на мышах, продолжительность жизни которых самая маленькая – 2 года. Причина ограниченной возможности потенциала их гипоталамуса связана с низкой степенью репараций, но высокой предрасположенностью к воспалению. Но многие животные живут 100 и более лет. Это означает, что темпы наступления провоспаления в гипоталамусе у них происходят на много медленнее за счёт высокой компенсаторной репаративной возможности. Очевидно механизмы провоспаления и определяют степень работоспособности гипоталамуса, а также будет ли животное относится к короткоживущим или к долгоживущим. Через эндогенный потенциал гипоталамуса природа регулирует продолжительность жизни. Как увядает гипоталамус, так увядает и весь организм. Но болезни организма в свою очередь тоже определяют темпы старения гипоталамуса. Как стареет организм, так стареет и гипоталамус. В данном случае это будут экзогенные факторы и условия, которые могут значительно варьировать и иметь переменчивые параметры. Тогда как темпы старения гипоталамуса обусловлены эндогенно, эволюционно зафиксированы в его возможностях. Почему этот потенциал обеспечивается через механизмы провоспаления, а вернее через способности или возможности клеток и ткани противостоять сенесценту?
Через гипоталамус происходит вся нагрузка онтогенетических перестроек, когда количественные изменения должны переходить в качественные. Все эти перестройки необходимо обеспечить сложнейшими механизмами компенсаций, чтобы при этом организм не вышел за пределы коридора гормезиса. От того как качественно и тонко проработаны обеспечительные механизмы этих перестроек зависит жизнеустойчивость этого органа.
Исчерпание, угасание его некоторых ресурсов у разных видов происходит по разному. Животные с длительной продолжительностью жизни получили в филогенезе более мощные ресурсы гипоталамуса, с повышенной устойчивостью к «скороспелости» и провоспалению. Возможно, это связано с более высокими функциональными возможностями гипоталамуса, более высокой работоспособностью, он медленнее изнашивается и меньше провоспаляется.
Очевидно, это связано большей развитостью этой структуры за счет заложения большего числа стволовых клеток, или более развитой системой ядер-регулировщиков, что усиливает его ресурс, не даёт так быстро воспаляться и изнашиваться. В таких гипоталамусах должны быть более развиты компенсаторы и тормозные механизмы, то есть система контрпозитных механизмов. Задача контрпозитов уравновесить систему так, чтобы не вывести её за пределы гормезиса.
Интенсивность воспалительной реакции гипоталамуса возрастает при всех возраст-зависимых заболеваниях, в том числе при метаболическом синдроме, раке… С гипоталамусом связаны гипертония, атеросклероз, климакс, гормональные дисфункции, доброкачественные опухоли, гиперплазии и весь «цвет» синдромокомплекса старения… При этом метаболический синдром, который приводит к диабету, избыточному весу и болезням сердца, появляется именно при воспалительных изменениях в гипоталамусе. Во всех случая как связанными со старением, так и с хроническими заболеваниями воспалительные сигнальные пути обычно сходятся к транскрипционному фактору NF-κB, одному из главнейших иммунных регуляторов. Чем больше этого белка в гипоталамусе мышей, тем быстрее у животного идут процессы старения.
Гипоталамус управляет системным старением посредством нейровоспалительной сигнализации.
Как стареет гипоталамус, а за ним вслед весь организм?
Учёные обнаружили небольшую группу клеток, управляющих старением. Ими оказались нейрональные стволовые клетки, известные своей способностью управлять формированием новых нейронов. Со временем численность нейрональных стволовых клеток в гипоталамусе сокращается, и этот факт ускоряет старение. Решили изучить судьбу этих клеток со временем у здоровых стареющих мышей. Оказалось, что численность стволовых клеток начинает снижаться примерно в 10-месячном возрасте, за несколько месяцев до появления первых признаков старения. Примерно к двум годам, когда проявляется старость, если речь идёт о мышах, — большая часть этих клеток уже исчезает.
В экспериментах если у мышей удавалось истребить стволовые клетки гипоталамуса, то происходило ускорение старения животного. Но в другом эксперименте достигли обратного эффекта по продлению жизни животного на 10%. Для этого им в гипоталамус ввели стволовые клетки, и их когнитивные и физические способности оставались в лучшем состоянии, чем у животных, которые не получали клеточных инъекций. Выяснили, что стволовые клетки в гипоталамусе выделяют огромное количество мембранных пузырьков – экзосом, наполненных разнообразными микрорегуляторными РНК (микроРНК). Так называют особые молекулы РНК очень небольшого размера, которые не участвуют в кодировании и синтезе белков, но могут вмешиваться в процесс трансляции, происходящей на других РНК. Однако, эффект от них был всего лишь 10%, то есть не существенный по сравнению с тем, что достигают животные голые землекопы, продолжительность жизни у которых до 27 лет, то есть увеличение более 1000%. Это является наводкой для построения концепции по решению проблемы отодвижения сенилита с помощью омоложения и восстановления функциональности гипоталамуса путём восстановления стволовых клеток гипоталамуса и уровня микроРНК в них.
Пузырьки с багажом из микроРНК можно получить из стволовых клеток, растущих в лабораторной культуре, в посуде с питательной культурой. И если эти пузырьки ввести в гипоталамус мышей, то эффект окажется тот же, что и при введении стволовых клеток. То есть стволовые клетки замедляют старение не только как ресурс клеток на замену, но и как источник неких молекулярных сигналов.
Ниже мы обсудим также и другой путь стимулирования репарации любых стволовых клеток, в том числе и гипоталамусе с помощью олигонуклеотидов, то есть строительных метаболитов РНК и ДНК. Данное направление намного проще по технологии, когда не надо ничего вводить в гипоталамус.
Гипоталамус не является главенствующей структурой действующей на принципах развития сам в себе, а работает на принципах обратной связи, когда динамично развиваются и зависят друг от друга низа и верха. Онтофизис (возрастные изменения) происходит одновременно и в гипоталамусе, и тканях мишенях. Омолаживать просто один гипоталамус бессмысленно или малоэффективно (на мышах это дало всего лишь 10% дополнения к жизни). Дело в том, что перезрелые ткани-мишени по системе обратной регулировки дадут быстрое нивелирование, исчезновение эффекта омоложения гипоталамуса.
Что общего между сенилитным провоспалением в гипоталамусе и болезнью Альцгеймера?
Болезнь Альцгеймера – это ускорение провоспалительного процесса в области мозга как одного из симптомов старения.
Выяснено, что интерлейкиновые провоспалительные вещества играют ведущую роль в запрограммированном старении не только мозга, но и всего организма.
Ведущей скрипкой в механизмах старения являются клеточные гормоны воспаления – интерлейкины. Интерлейкины в малых количествах приносят пользу организму, а в крайних случаях при их избытке – огромный вред. При старении они уже выступают не как гормоны противодействия, а ключевым инструментом, гормонами смерти, которые выключают наш организм, отключают жизнедеятельность его важнейших органов: сердце, лёгкие, мозг… Это авральная программа на ускорение самоуничтожения, самоликвидации организма. Как результат именно последствий воспалительных интерлейкинов – гибель организма. Они являются важнейшей причиной мортального = смертельного исхода при всей неизлечимой хронике. Это механизм, задуманный природой по отключению организма, по частям отключения его важнейших систем жизнедеятельности. Это механизм завершения, отключения, несовместимости ситуации с дальнейшим пролонгированием жизни. По сути, его можно сравнить с механизмом активного апоптоза, но на уровне всего организма = феноптоза – механизма самоуничтожения. Дальше нет смысла поддерживать этот разваливаюшийся организм как систему систем. Это механизм, обеспечивающий мортальность для хронических безвыходных ситуаций. Эволюция не направлена на поддерживание существования больных, старых и изнемогающих организмов. Это специальный механизм отключения, элиминации. Всё тяжело больное должно быть элиминировано!
Доказано, что такой же механизм провоспаления имеется и при старении организма. И в том и в другом случае организм запускает механизм самоликвидации, апоптоза на уровне всего организма. Оно завязано с онтогенетической программой, то есть «протягивания» организма с одной фазы, этапа развития до другой, и, в конечном итоге, на фазе сенилита прорываются «шлюзы» по переполнению организма провоспалительными веществами, которые и обеспечивают ускорение лимитирования жизни. Механизмы противостояния провоспалениям исчерпаны. Интерлейкины обеспечивают фон, почву, на которой обязательно разовьются, вспыхнут буйным цветом мортальные болезни старости - анизомалии.
В тоже время, выше, мы приводили данные, что у мышей максимальные уровни провоспалительных гормонов отмечены в гипоталамусе. Значит у мышей он определяет продолжительность жизни этого вида животных. У животных долгожителей, явно гипоталамус не будет ведущей скрипкой для определения сроков феноптоза, а эту роль возьмут на себя другие ткани, в том числе и ткани мозга, сосудов и др… Сроки феноптоза будут уже в большей степени определяться не гипергипоталамическим доминированием, а за счет стем-доминировния в тканях.
Преодоление провоспалительного барьера в борьбе с болезнями старости является важнейшей составной задачей комплексных лечебных мероприятий.
Любое провоспаление всегда сопряжено с:
- оксидативным стрессом,
- митохондриальной дисфункцией,
- снижением уровня антиоксиданта Глутатиона,
- эпигенезом.
Сенилитное провоспаление можно рассматривать как следствие неизбежных программ, которые включаются на определённых этапах онтогенеза, а можно рассматривать, что провоспаление это не самостоятельный процесс, а всего лишь следствие указанных процессов? Кто из них первичен? Однозначно можно утверждать, что перечисленные процессы – это всего лишь механизмы реализации провоспаления и сенесцента.
В качестве веществ, противодействующих провоспалительным процессам, являются Омега-3 кислоты, Аспирин, Куркумин-Адванс, витамин D3, Мелатонин...
Можно ли полностью погасить сенилитные провоспаления? Да, можно, но для этого нужно понять как можно полностью репарировать все точки самообновления, самоомоложения.
Биохимическая возрастная динамика гипоталамуса
Найдено вещество менин, которое играет важную роль в подавлении нейровоспаления и развитии мозга. Показано, что скорость старения гипоталамуса сопряжена с уровнем снижения белка менина, который производится в головном мозге и участвует в регуляции нейровоспаления.
Восстановление экспрессии менина в вентромедиальном ядре гипоталамуса (VMH) у пожилых мышей увеличивало продолжительность жизни, улучшало обучение и память, и улучшало биомаркеры старения, в то время как ингибирование менина в VMH у мышей среднего возраста вызывало преждевременное старение и ускоряло снижение когнитивных способностей.
Снижение уровня менина в гипоталамусе приводило к когнитивным нарушениям, возрастным изменения в костной массе и толщине кожи, а также умеренное сокращение продолжительности жизни мышей.
Итак, уровень менина в гипоталамусе, но не в астроцитах или микроглии, снижается с возрастом. Чтобы проверить, можно ли обратить процесс вспять и вернуть нарушенные функции, исследователи доставили ген производства менина в гипоталамус пожилых (20-месячных) мышей. 30 дней спустя они обнаружили уменьшение толщины кожи и изменение структуры костной массы, а также улучшение обучения, познания и равновесия. Изменения коррелировали с увеличением уровня D-серина в гиппокампе.
Аналогичные изменения в работе когнитивных функций наблюдались также после трёх недель приёма подопытными мышами пищевых добавок с D-серином. Но препарат не у всех проходит через гематоэнцефалический барьер, поэтому более лучше подходит Саркозин. Саркозин считается более надежной альтернативой. D-серин является ко-агонистом рецепторов NDMA, то есть усиливает действие других химических соединений (в частности, глутамата и N-метил-D-аспартата), имеющих отношение к этим рецепторам. D-серин часто относят к категории ноотропных средств.
Дозировка D-серина. Обычно, фигурирует 30 мг/кг массы тела. Но удвоение или учетверение стандартной дозировки до 60 мг/кг и 120 мг/кг, соответственно, усиливает.
Передозировка. Передозировка данной аминокислотой практически не возможна, нет данных о случаях передозировки Серином.
Взаимодействие. Вещество отлично сочетается с прочими лек. средствами.
Элевационные и инволюционные изменения в гипоталамусе
Выяснено, что у людей среднего возраста с избыточным весом гипоталамус заметно больше, чем у тех, кто имеет нормальный вес, но того же возраста. Также у людей среднего возраста гипоталамус имеет увеличенные размеры (гипертрофия) по сравнению с нормальными размерами у людей более молодого возраста, а в старости он уменьшается. Очевидно всегда существует связь между гипертрофией гипоталамуса и так называемым воспалением гипоталамуса, а также его гиперфункцией, перезреванием. В всех случаях это аномальное состояние. Организму они не выгодны. Эти процессы находятся за пределами оптимума функционирования системы или органа и при продолжении ведут к его истощению, ослаблению. Подчеркиваю, что речь идёт именно о воспалении мозга, а не клеточном воспалении или на уровне иммунных реакций. Показано, что при хроническом переедании, питании без значительных перерывов в первую очередь страдает гипоталамус. Помочь в этом случае может интервальное питание с ограничением калорий.
«Изнашивание» = перезревание гипоталамуса, в том числе и гипоталамическая возрастная инволюция, онтогенетические трансформации могут происходить от многочисленных причин, в том числе при хронических стрессах, болезнях… Преждевременный климакс и преждевременное старение тоже обеспечиваются через механизмы гипоталамуса.
Продление жизни и неотения (удлинение периода молодости) тоже опосредуются через гипоталамус. Гипоталамус можно расценивать как водитель ритмов = пейсмейкер, где самый большой ритм – длительность жизни. Но изнашивается и стареет гипоталамус и от состояния гормональной перезрелости, избыточности, затем и климакса. Зрелость и перезрелость пролог к старению, а точнее неизбежные последовательные этапы онтогенеза адальтус → матура → сенилит. Одно ведёт к наступлению другого. Климакс при этом следует рассматривать как одно из проявлений резистентности, нечувствительности данного регулирующего органа к различным сигналам нервной системы, к гормонам, нейромедиаторам и др. факторам. Это проявляется как этап истощения, исчерпания и выход за пределы возможности после длительной гиперфункции. После длительной гиперфункции всегда наступает откат в виде гипофункции; после гипертрофии → атрофия. Это приобретение возрастной нечувствительности, толерантности. Регулирующая система при этом выходит за пределы своего оптимума, то есть за пределы коридора гормезиса и работает не на полную мощь, а в итоге вразнос.
Движущие силы онтогенетических изменений в гипоталамусе можно разделить на два уровня:
- Нисходящие процессы с верхних этажей пирамиды систем организма, в частности верховного регулятора, в котором происходит вынужденная элевация (наращивание мощи, гиперфункции, гипертрофии, перезрелости) механизмов верховных гормональных регулировок. Они ведут к гипертрофии и гиперфункции.
- Восходящие с клеточного уровня, которые обусловлены уменьшением количества и ослаблением возможностей стволовых клеток и экзосомальных миРНК с превалированием клеток прогениторного типа, то есть проявления феномена так называемого соматического доминирования, когда состояние ткани и её потребности в репарации определяют не стволовые, а соматические клетки, что, очевидно, связано с лимитом Хейфлика, то есть репликативной ограниченностью клеток. Они ведут к гипофункции и атрофии.
Таким образом, можно утверждать, что гипоталамус стареет в два этапа:
1-й этап гиперфункций – ведёт к адальтусу* и матуре*;
2-й этап гипофункций – ведёт к сенесценту* + сенилиту* = геронтос*.
Первый этап сопряжен с элевационными процессами по Дильману. Второй этап сопряжен с угасанием, инволюцией органа по Гращенкову. Таким образом, по Гарбузову геронтос – это двухэтапный процесс старения от элевации до инволюции. Кроме того, геронтос – двусторонний процесс: от сенилита до сенесцента. Особенностью данной гипотезы является то, что в процесс общего старения включен еще и сенесцент. Таким образом, геронтос - это единство сенесцента и сенилита, то есть взаимоувязанных процессов. Причём сенесцент вторичен, навязан и не является первопричиной геронтоса, а наоборот следствием геронтоса, также сенесцент не является неизбежным следствием лимита Хейфлика.
При этом Гарбузов показывает, что сенесцент это проявление, следствие онтофизиса*, а не формального проявления лимита Хейфлика, феномен который связывали с укорочением теломер. Причем сенесцент и онтофизис являются следствием эпигенетических перестроек, в отличие от всех остальных этапов онтогенеза, которые являются программированным морфогенезом.
Такое наращивание происходит из-за постепенного приобретения резистентности и толерантности в данной системе с обратными связями: верховный регулятор – железы и органы мишени. С возрастом большое количество функций ослабевает из-за появления в системе звеньев с резистентностью, они попросту недорабатывают, ослабевают. Резко возрастают на стадии зрелости = матуры количество половых стероидов как тестостерон, эстрогены, а затем их количество идёт на убыль. С другой стороны уменьшается количество выработки мелатонина, гормона роста - соматропина и др. Меняется баланс нейромедиаторов, в частности увеличивается, а затем уменьшается кортизол, снижается ацетилхолин и т.д. Вся система регулировок сдвигается, причем не каждый отдел сдвигается сам по себе по своим правилам, а в соподчинении с другими параметрами гормонов. Такие вторичные программы и механизмы соподчинения предусмотрены генетически в филогенезе. Весь этот комплекс разноплановых систем регулировок, которые движутся в разных направлениях, есть проявление общего сдвига регулировок.
Клеточное старение (сенесцент) как проявление клеточной нечувствительности, неуправляемости и перехода на эпигенетическое регулирование, когда оно становится доминантным началом в жизненных циклах клетки
Напомню, что сутью сенесцентных клеток является как репликативное торможение, так и усиление в них нечувствительности, толерантности во многих сигналинговых путях и рецепторных структурах, что ведёт клетки к рецессии, то есть снижению их как функциональной работоспособности, так и активность стволовых клеток. Такие клеточные системы начинают вырождаться. Подробно об этом в моей книге: «ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ» [2]. Для того чтобы объяснить природу сенесцента клеток, следует предварительно рассмотреть как клетки входят в состояние стресса, а затем в состояние сверхадаптации. Речь пойдет именно о клеточном стрессе, а не общеорганизменном, известного под названием Общий Адаптационный Синдром (ОАС), который описал Г. Селье. Факторов клеточного стресса бывает множество, но механизмы их стресса и итоговые последствия тоже универсальны, сходны. Автором предложено их обозначить как Адаптационный Универсальный Клеточный Ответ (АУКО). Он может быть условным, то есть вызванным внешними факторами, так и безусловным, то есть врожденным, связанным с внутренними факторами онтогенеза и онтофизиса. В связи с этим сенесцент следует рассматривать тоже как следствие внутренних факторов условий развития. В разделе: Гормезиорез и гомеорез – причина сенесцента, сенилита через эпигенез и деградацию белков показано, что причиной клеточного хронического стресса = сенесцента может быть как выход организма в целом за пределы гормезиса, связанный с возрастом, так и вычленение и содержание клеток вне организма в условиях культуры ткани, когда всё это выводит на длительное функционирование клеток за пределами гомеостаза. В итоге жизнедеятельность таких клеток начинает выходить за пределы оптимума. Как результат снижается ВИТАУКТ* организма и Жизненная Сила клеток. Механизмы клеточного стресса реализуются через образование белков теплового шока (БТШ), которые запускают белки-сиртуины, в частности SIRT1, вслед за которыми начинаются эпигенетические перестройки со многими последствиями. Любой стресс активирует клеточные функции, делает их более чувствительными за счёт механизмов АУКО*. АОС начинается с активации надпочечников и усиливает выработку ими гормонов стресса адреналина, кортизола, глюкокортикоидов, обладающих способностью стабилизировать мембраны, другие гормоны работают как экспрессоры для генов. Клетка приобретает повышенную сопротивляемость к негативным факторам, функциональность, но при этом может выйти за пределы гомеостазов, а значит работает на износ. К внутренним негативным факторам можно относить и онтогенетический ответ клеток на возрастные изменения в организме. Причем в отличие от более ранних этапов онтогенеза, вплоть до юности, они сопровождаются генетическими регулировками, включая механизмы морфогенеза, например пубертатное оволоснение. Но в последующем динамика управления онтогенезом с переходами на этапы адальтуса, матуры, сенилита переходит на превалирование регулирования на уровне эпигенеза. Эпигенез становится доминантным фактором, «жокеем» дальнейшего развития с целым шлейфом различных анизомалий. Он же эпигенез становится и ограничительным фактором, обуславливающим мортус, то есть феноптоз*. Этим тип феноптоза человека отличается от рыбы горбуши, причиной смерти которой является выраженное гипергипоталамическое доминирование, ведущего к избытку репродуктивных гормонов с многочисленными последствиями не совместимыми с жизнью.
В любом случае это режим в одних условиях для дегенеративных перерождений, утрате тканью дифференциальных функций, а в других – путь к сенесценту или онкогенезу. Особенность клеток животных в том, что к сенесценту клетки изначально предрасположены, настроены на это. Но это не программативная настроенность, а коммитативная – то есть на уровне эпигенеза – многогранный комплекс изменений происходящий, как в состоянии стресса, так и онтофизиса.
Как гипоталамус будучи гетерогенной, многофункциональной структурой определяет сенилит
За что отвечают ядра гипоталамуса?
В ядрах гипоталамуса происходит тончайшая координация деятельности вегетативной нервной системы, которая управляет всеми внутренними органами, регулирует процессы обмена веществ в организме.
Благодаря четкой и слаженной работе различных отделов гипоталамуса сохраняется относительная устойчивость различных функций организма, что совершенно необходимо для его нормального существования.
За что отвечает гипофиз и гипоталамус?
Гипоталамо-гипофизарная система регулирует такие важные функции организма, как температура тела, сон, аппетит, обмен веществ, половое развитие, репродуктивную функцию и многое другое. Она также контролирует выработку таких гормонов, как гормон роста, пролактин, АКТГ, тиреотропин, лейтенизирующий гормон и фолликулостимулирующий гормон.
Гипоталамус. Характеристика основных ядерных групп. Участие гипоталамуса в регуляции вегетативных функций и в формировании эмоций и мотиваций
Группа передних ядер. Это супраоптическое, супрахиазматическое, паравентрикулярное и др.
Средняя группа ядер. Это вентромедиальное, дорсомедиальное и др.
Наружная группа ядер. Образована латеральным гипоталамическим ядром, ядром серого бугра и т.д.
Задняя группа ядер. Состоит из заднего гипоталамического, перифорникального, премамиллярного, медиального мамиллярного и латерального мамиллярного ядер и др.
Особенностью гипоталамуса является то, что его ядра могут возбуждаться двумя способами. Первый способ – это нервный путь. Второй способ – это возбуждение ядер гипоталамуса гуморальными влияниями. Данная особенность гипоталамуса связана с наличием в гипоталамусе 3 типов нейронов. Первый – нейросекреторный. Второй тип нейронов – ретикулярный. Третий тип нейронов – это слабоветвящиеся нейроны с короткими дендритами.
Связи гипоталамуса. Он имеет хорошо развитые афферентные и эфферентные пути. Он получает афферентные пути из коры больших полушарий мозга, из таламических ядер, ядер базальных ганглиев, ретикулярной формации среднего мозга.
Ядра переднего гипоталамуса – супраоптическое и паравентрикулярное – связаны системой волокон с гипофизом. Эти волокна служат не только для проведения импульсов, но и для транспорта продуктов нейросекреции, которые вырабатываются нейронами этих ядер (вазопрессин, окситоцин, статины, либерины).
В составе гипоталамуса находятся нейроны, вырабатывающие гормоны или специальные вещества, которые в дальнейшем, действуя на клетки соответствующих эндокринных желез, приводят к выделению или прекращению выделения гормонов (так называемые рилизинг-факторы). Все эти вещества вырабатываются в нейронах гипоталамуса, затем транспортируются по их аксонам в гипофиз. Ядра гипоталамуса связаны с гипофизом гипоталамо-гипофизарным трактом, который состоит примерно из 200 000 волокон. Свойство нейронов вырабатывать специальные белковые секреты и затем их транспортировать для выброса в кровяное русло называется нейрокринией.
Гипоталамус является частью промежуточного мозга и одновременно эндокринным органом. В определенных его участках осуществляется трансформация нервных импульсов в эндокринный процесс. Крупные нейроны переднего гипоталамуса образуют вазопрессин (супраоптическое ядро) и окситоцин (паравентрикулярное ядро). В других областях гипоталамуса образуются рилизинг-факторы. Одни из этих факторов играют роль гипофизарных стимуляторов (либерины), другие — ингибиторов (статины). В дополнение к тем нейронам, аксоны которых проецируются в гипофиз или в портальную систему гипофиза, другие нейроны этого же ядра отдают аксоны в многие участки головного мозга. Таким образом, один и тот же гипоталамический нейропептид может выполнять роль нейрогормона и медиатора или модулятора синаптической передачи.
Велика роль в регуляции и координации функций вегетативной нервной системы. В регуляции функции её симпатической части участвуют ядра задней области гипоталамуса, а функции парасимпатической части вегетативной нервной системы регулируют ядра его передней и средней областей. Стимуляция передней и средней областей вызывает реакции, характерные для парасимпатической нервной системы — урежение сердцебиений, усиление перистальтики кишечника, повышение тонуса мочевого пузыря и др., а раздражение задней области проявляется усилением симпатических реакций — учащением сердцебиений и т.д.
С состоянием вегетативной нервной системы тесно связаны вазомоторные реакции гипоталамического происхождения. Различные виды артериальной гипертензии, развивающиеся после стимуляции Г., обусловлены комбинированным влиянием симпатической части вегетативной нервной системы и выделением адреналина надпочечниками, хотя в данном случае нельзя исключить влияние нейрогипофиза, особенно в генезе устойчивой артериальной гипертензии. Таким образом, возрастную гипертонию, хроническую усталость, бессонницу… можно рассматривать не только как примеры разболтанности, изношенности, разрегулировки неврологической стороны деятельности гипталамуса, но и как возрастных неизбежных его перестроек, связанных с онтогенезом. Тогда как поседение волос, ухудшение зрения, ухудшение качества кожи, морщины на ней, мышечная саркопения, атеросклероз, диабет… – проявления трофической функции гипоталамуса в целом и в частности его ядер определяющих это. Гипоталамус стареет гетерохронно и гетерогенно. Все многочисленные ядра являются представительствами многочисленных функций регулировок, за которые они отвечают. У каждой функции, системы имеется своё ядро-представительство в Генштабе. Деятельность всех этих представительств согласована. С перестройками в одних системах могут происходить подстройки в других. Обобщенно - весь этот спектр возрастных изменений деятельности гипоталамуса – это многочисленные проявления выхода за границы коридора Гормезиса, когда происходит потеря многочисленных сил Витаукта – интегральной Жизненной Силы. Обобщенно - Витаукт это сила определяющая слаженный интегрированный оптимум работы всех систем. Это сила, а точнее программа гармонизирующая работу всего Генштаба. Витаукт состоит из многочисленных ручейков автоматической саморегулировки, когда их оркестр играет одну симфонию под названием Гармония.
Итак, гипоталамус гетерогенная структура, развивающаяся гетерохронно и, тем не менее, определяющая специфику сенилита, стадий онтогенеза. Скорость онтогенеза определяют не все ядра гипоталамуса, а только те, которые определяют репродуктивную зрелость и трофику тканей.
К числу гипоталамических ядер, ответственных за реализацию программы старения, предположительно относят супрахиазмальное ядро и срединную группу ядер, включая аркуатное, вентромедиальное и дорсомедиальное ядра. Процесс старения в гипоталамусе сопровождается:
- уменьшением частоты импульсации нейронов,
- нарушением обмена кальция, магния,
- увеличением продукции оксида азота,
- снижением аутофагии,
- нарастанием нечувствительности, рефрактерности к сигналам (невосприимчивости), толерантности,
- усилением првоспалительных процессов,
- оксидативный и нитрозативный стресс,
- снижением уровня эргичности клеток, в т.ч. выработки АТФ,
- нарушением процессов внутриклеточного сигналинга, в частности Sirt1, mTOR, NF-κB, AMPK, P53, FoxO…
- …
Нейроны ДЯ и ДМЯ содержат медиаторы глутамат и ГАМК, при этом доля последнего преобладает. В ВМЯ преобладают глутаматергические нейроны, а доля ГАМК-ергических невелика. При старении у крыс усиливаются взаимно противоположные процессы возбуждения и торможения, по-видимому, нарушая баланс ГАМКергической и глутаматергической систем.
Также важная роль в гипоталамическом старении принадлежит таницитам, что выражается в нарушении их связи с нейронами и барьерной функции. Стволовые клетки гипоталамуса, расположенные преимущественно в области срединной группы ядер, выделяют экзосомальные миРНК, которые также ответственны за антивозрастные эффекты этих клеток. При этом количество самих стволовых гипоталамических клеток и экзосомальных миРНК уменьшается по мере старения.
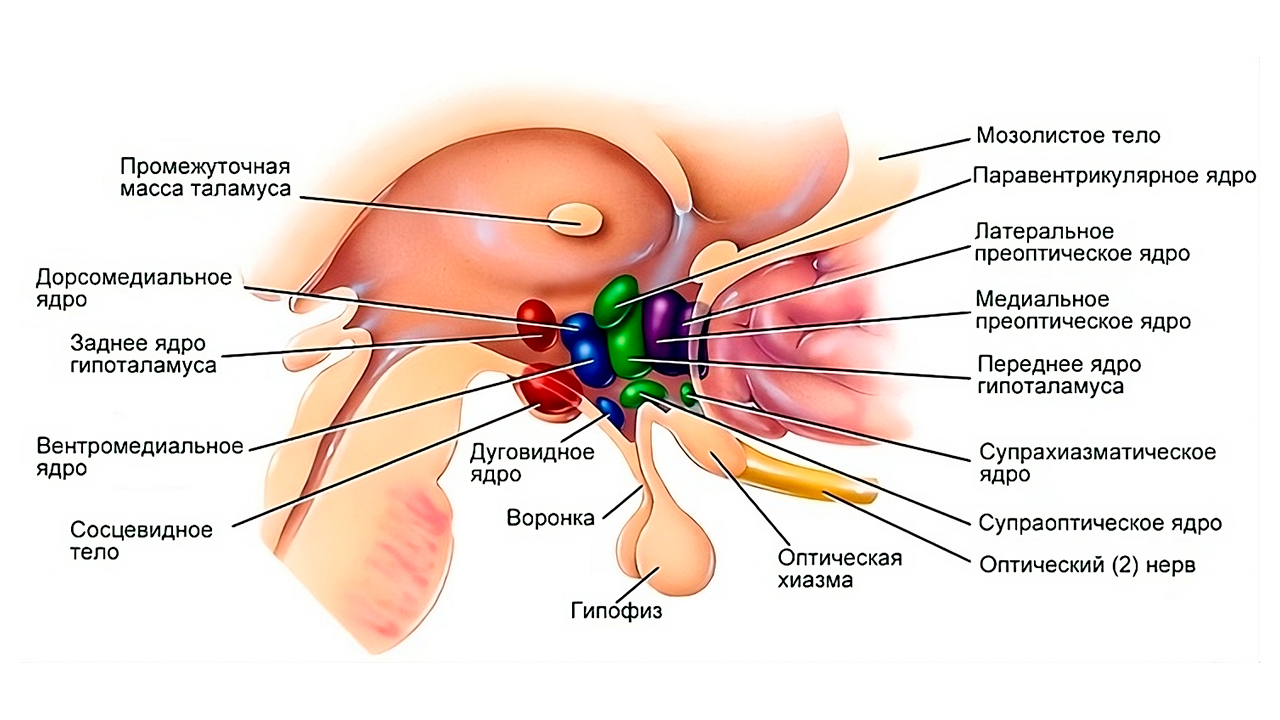
Рис. 1. Многоядерное строение гипоталамуса. Гипоталамус как орган отвечает за многочисленные нейрогормональные функции, в том числе и за созревание организма и его старение.
Важным отличием и особенностью гипоталамуса животных относящихся к долгожителям является наличие в гипоталамусе демфинговых структур (амортизаторных), механизмов, которые сглаживают провоспалительные процессы на уровне клеток, тем самым пролонгируя функционал этих клеток. Более мощные и самоомолаживающиеся гипоталамусы обеспечивают более продвинутые сроки продолжительности жизни.
Способность к самоомолаживанию гипоталамуса обеспечена на коммитационном уровне. Животные долгожители коммитированы на мощное самоомоложение, реювенилизацию клеток гипоталамуса, преодоление в них сенесцента. Это аналогично тому как жестко регулируется уровнем гормонов и их соотношением продолжительность жизни 1- или 2-х летних растений, в отличие от их антиподов многолетников. Уровень гормонов реювенилизаторов – цитокининов у короткоживущих растений практически отсутствует, не предусмотрен, тогда как у долгоживущих такая возможность выработки реювенилизаторов постоянно открыта, что обеспечивает закладку новых почек для роста. В свою очередь у короткоживущих животных не предусмотрены демферные механизмы для сдерживания и противодействия провоспалению. Регулировка происходит через посредство главного апекса – аналога гипофиза. Продолжительность жизни короткоживущих растений коммитирована специальными механизмами. К такому же типу регулировок продолжительности жизни относится и рыба горбуша, которой строго предписана монокарпическая жизнь, то есть отмирание после первого нереста в 8 лет. Этот тип регулировки онтогенеза животных определяется как гипергипоталамическое доминирование, а у растений 1- и 2-х леток как гиперапикальное доминирование.
Можно ли остановить возрастное саморазрушение гипоталамуса и остановить старение?
Также обнаружено, что воздействие этой потери стволовых клеток обратимо. Восстановление клеточного пула или поступление молекул, вырабатываемых этими клетками, способно замедлить процесс старения и даже обратить вспять многие из его эффектов. Это показано в экспериментах доктора Цая. Важно было для понимания природы старения гипоталамуса ответить на вопрос является ли потеря нейрональных стволовых клеток причиной старения или просто идёт параллельно с ним. Для этого учёные выборочно разрушили стволовые клетки гипоталамуса у мыши среднего возраста. Это ускорило процессы старения по сравнению с животными из контрольной группы. Кроме того, мыши, клетки которых были уничтожены, скончались раньше, чем обычно.
Важно было получить ответ и на противоположный вопрос: можно ли путём добавления клеток в гипоталамус противостоять старению? Для этого они пересадили стволовые клетки как мышам, собственные клетки которых были уничтожены, так и обычным мышам, не подвергавшимся специальным процедурам. Лечение замедлило или обратило вспять процесс старения у животных из обеих групп. Обнаружено, что «антивозрастной» эффект нейрональных стволовых клеток объясняется их способностью вырабатывать микроРНК. Этот тип РНК не участвует в синтезе белков, но играет ключевую роль в управлении экспрессией генов. МикроРНК была упакована в небольшие частицы, экзосомы, которые выделялись в спинномозговую жидкость мышей. Исследователи выделили экзосомы из клеток и впрыснули их в спинномозговую жидкость двух групп мышей. Все животные были среднего возраста, а группы отличались тем же, чем и в предыдущих экспериментах — у части животных стволовые клетки гипоталамуса были избирательно уничтожены. В обеих группах инъекция микроРНК замедлила старение — это было продемонстрировано при помощи анализа тканей и проведения специальных тестов поведения. В ходе тестов учёные оценивали мышечную выносливость, координацию движений, социальное поведение и способности к обучению.
Возможности аутофагии в омоложении гипоталамуса
Стало понятно, что старение гипоталамуса сопряжено с мощным провоспалительным процессом в нём с последующей гипертрофией, а затем дегенерацией и атрофией его. Гипоталамус больше всех мозговых тканей страдает от провоспаления. В свою очередь стало известно, что стимулирование процесса аутофагии может стать мощным оружием в борьбе с провоспаление. Подробности можно прочитать в статье Гарбузова Г.А.: «Омоложение через аутофагию. Интервальное питание – путь к идеальному здоровью».
Кроме борьбы с провоспалением аутофагия проявляет свойства сенолитика против сенесцентных клеток. В последние годы всё чаще обращают внимание на сенолитики, вещества которые способствуют вытеснению стареющих линий клеток. Напомню, что аутофагия направлена на борьбу с клетками перегруженными отходами, что также имеется и стареющих клеток. А ведь сенолитизм (самоуничтожение стареющих клеток) это один из возможных вариантов аутофагии. Считается, что с возрастом число сенесцентных клеток в ткани увеличивается и это является причиной её старения. Очевидно где сильнее проявляется провоспаление, там и сильнее процветает сенесцент клеток.
Следует понимать, что любое воспаление состоит из двух противоположно действующих начал: оно может повести по пути отмирания, в первую очередь высоко функциональных клеток (дегенерация), или инициации защиты в виде роста и обновления клеток (самоомоложение). Это тонкий маятник саморегулировки, который нужно уметь правильно направить в нужном направлении. Очевидно мягкие аутофагические процессы в виде микроголодания, то есть одноразового питания - лучшее средство-камертон в самонастройке этого маятника.
Возможности омоложения гипоталамуса путём повышения уровня выработки мелатонина эпифизом
Подробности об этом можно прочитать в статье Гарбузова Г.А.: «ГИПОСЕНСОРНАЯ КАМЕРА (камера флоатинга)» garbuzov.org [5].
Нами предложено использовать эту камеру для погружения в состояние заглубинного сна для снятия провоспалительных последствий и сцепленных с ними возраст-зависимых заболеваний, включая и раковые.
Только в этой камере реально осуществить процесс, который связан с неким тета-ритмом, погружения мозга в глубочайший покой. Это фаза глубокого расслабления мозга, выходящего за пределы расслабления в обычном сне. Только так снимаются все ограничения по дозированной ограниченной выработке мелатонина. Его выработка переходит из циркадного ритмичного производства в непрерывное, причем в огромных количествах лавинообразно. Только в таких условиях возможно подавить хронический пожар провоспаления.
Становится очевидным, что организм борется как с сенесцентными-провоспаленными клетками, так и с опухолевыми не за счёт иммунитета, а путём «смывания» их программ. Под термином «смывание» подразумевается разблокирование, экспрессия более древних первичных генов, несущих контрпозитные функции на репарацию, регенерацию или блокировку аномалий. Мелатонин просто расклинивает «заклинившие» клеточным программы гликолиза, выводит их из ступора.
Приведенные случаи исцеления явно завязаны на противостояние механизмам провоспаления и запуска механизмов апоптоза и регенерации в опухоли.
Лечение в гипосенсорной камере в сочетании с лечебным голоданием лучшее новационное решение для разблокировки онко- и сенесцентных программ. Дело в том, что онкология и сенесцент клеток – это не разные процессы, а два крайних состояния одного общего процесса выхода за пределы коридора гомеостаза клетки в условиях гликолиза, а другими словами это два разных берега одной и той же проблемы – анаэробного метаболизма. В обоих случаях запускаются мощные механизмы провоспаления, когда в одном случае клетки выходят за рамки гомеостаза на путь неограниченного деления, а в другом случае наоборот происходит полное торможение репликационных механизмов. Погашение провоспалительных механизмов в обоих случаях приведёт систему в русло гомеостаза. Причём задача вернуть в русло Витаукта параметры как гомеостазов, так и гормезиса. В ряде случаев одно цепляется за другое.
Гипоталамус в роли осцилятора темпов продвижения по этапам онтогенеза или участник в генезе эндогенного времени, то есть обуславливает продолжительность жизни вида
Выше мы рассматривали вопрос о разной продолжительности жизни у разных видов животных, например мыши живут 2 года, голый землекоп – 27 лет, собака - 20, галапогосская черепаха – 200 лет и т. д. Даже внутри вида возможна существенное различие продолжительности жизни: так у пчел рабочие особи живут 56 дней, а матки – 3-5 лет. То есть на их жизненный цикл сказывается не генетические изначальные клеточные программы, а верхние регуляторы. Причем обычная трудовая пчела может при необходимости стать маткой. Темпы старения и онтогенеза определяют возможности гипоталамуса. В связи с этим некоторые авторы предложили ввести понятие о гипоталамусе как регулятора эндогенного времени = Тэнд. Если у каждого гомеостаза есть всегда показатель его constanta, то у гормезиса этим показателем будет Тэнд.
Неотения сдерживает сенилит через гипоталамус
Сенилит или верхний уровень обусловленности старения можно сдерживать и отодвигать только преодолевая механизмы гиперфункциональности = перезрелости и климакса. Гиперфункциональность ведёт к нечувствительности = резистентности. Реализовать сдерживание её можно только за счёт подключения механизмов неотении, которая обеспечит сдерживание элевационных процессов нисходящих из гипоталамуса. Это означает обеспечение молодости низов обеспечит молодость верхов, и наоборот обеспечение молодости верхов обеспечит молодость низов. Низы определяют верха, а верха определяют низы. Без неотении мыши живут 2-3 года, а с неотенией голые землекопы – 30 лет. Но для продления неотении, продвижении её во времени нужны обеспечивающие это механизмы. Но откуда идёт первичная динамика изменений: сверху или снизу? С чем это связано, откуда такая предрасположенность: с особенностями замедленного созревания низов или особенностями их гипоталамуса, который медленнее созревает?
И это связано с особенностями их гипоталамуса, этапы зрелости и климакса в гипоталамусе у них наступают намного позже. Причем неотения может быть врожденной, обеспеченной обязательством клеточных программ, а может быть искусственной, приобретённой за счёт сочетания определённого ряда факторов.
У рыбы горбуши этап старения начинается в девять лет при их нересте, когда гипоталамус работает вразнос и происходит резкий скачок изменений гормонального профиля в виде гормонального перезревания и чрезмерной работы гипофиза. Гонады являются железой-мишенью гипоталамуса и при их удалении происходит предотвращение нереста и гибели тихоокеанского лосося нерки: рыбы живут в 2-3 раза дольше и умирают при явлениях постепенного старения. В данном случае эту особенность старения горбуши можно расценить как разновидность прогерии – одного из типов преждевременного старения. По сути, прогерия противоположность неотении. Гипоталамус лишь заводит темпы старения, перезревания во всех подведомственных системах, но не является прямым фактором старения. Его можно назвать прибором осцилирующим инициатором-модулятором темпов старения. У разных видов этот осцилятор темпов продвижения по этапам онтогенеза работает по разному. Всё зависит от того как он настроен в филогенезе. От особенностей гипоталамуса зависят темпы прогерии или неотении.
У человека, тем не менее, удаление гонад незначительно влияет на увеличение его продолжительности жизни, всего на 14-17 лет. Очевидно кастрирование в детском возрасте несколько больше сдерживает перезревание и старение гипоталамуса. Но у них начинаются другие проблемы, в том числе ожирение, остеопороз, гинекомастия… Кастрация баранов увеличивает их продолжительность жизни на 60%.

Рис. 2. Рыба горбуша, где нижний экземпляр находится на стадии крайней матуры и сенилита.
У других типов лососевых рыб такой строгой детерминации старения нет и это тоже связано с особенностями быстрого или замедленного созревания гипоталамуса. Его возможности сильно определяют темпы наступления старения животных.
Выше мы говорили о гипоталамическом синдроме, который ведёт к хроническим многим заболеваниям и ускоренному старению.
Но практике известен и противоположный синдром – гипофизарный нанизм, когда происходит частичное недоразвитие гипоталамуса и тормозятся ряд его центров, когда недоразвиты гонады и гормоны роста. Напомню, что у горбуши происходит гипергонадный кризис и перезревание. У животных неотеников происходит обратный процесс – отставание, замедление полового созревания, например у голого землекопа.

Рис. 3. Мужчина 32 лет, который выглядит как 14-летний школьник. Все признаки неотении по причине гипофизарного нанизма (недоразвития гипоталамуса).
Типичным примером неотении является случай с 32-летним мужчиной, который перестал взрослеть-стареть 20 лет назад и выглядит как 14-летний школьник. Зовут его Денис Вашурин. Ясно одно, что такая аномалия его развития в виде неотении, точно также как и у животного голого землекопа увеличила продолжительность жизни в 10 раз, увеличит и у этого человека его продолжительность жизни значительно, возможно кратно.
Предлагаю познакомиться со статьей Гарбузова Г.А.: «НЕОТЕНИЯ – ПРОДЛЁННАЯ МОЛОДОСТЬ, МОЖНО ЛИ ЕЁ СОХРАНЯТЬ НЕОГРАНИЧЕННО С ПОМОЩЬЮ АУТОФАГИИ?» на сайте garbuzov.org.
Возможности интервального голодания в сохранении гипоталамуса
Продление неотении – единственный способ поддерживать юность и отодвинуть старение. Это возможно только через сохранение постоянства и репарации в гипоталамусе. Задача найти ключи к его управлению. Частое питание, а точнее трёхразовое, да ещё и с перекусами, является стрессом и перегрузкой для гипоталамуса, рано или поздно это проявится в его истощении и изнашивании. «Полетит» вся система контроля. Гипоталамус работает по принципу «камертона» - настраивает и поддерживает правильный тон оркестра. Это уже «разболтанный прибор», который работает вразнос, в диссонансе и его надо восстанавливать, успокоить...
Разработка способов модуляция гипоталамической аутофагии позволит дать ключи к различным терапевтическим вмешательствам для замедления старения. Одним из таких ключей является аутофагия, о которой можно прочитать в статье Гарбузова Г.А.: ОДНОРАЗОВОЕ ПИТАНИЕ ДЛЯ ДОСТИЖЕНИЯ АУТОФАГИИ КАК ПУТЬ ОМОЛОЖЕНИЯ И ДОСТИЖЕНИЯ ИДЕАЛЬНОГО ЗДОРОВЬЯ на сайте garbuzov.org.
Дело в том, что первым страдает гипоталамус при хроническом питании без интервалов голода нужных для аутофагии. В эксперименте мыши жили дольше именно те, которые сочетали не только ограничение калорий в пище, но и были переведены на интервальное питание.
Когда вы придерживаетесь популярной схемы интервального голодания, не употребляя пищу 8 часов до и после полуночи, то гонадотропин-рилизинг-фактор начинает работать активнее и воспаление в мозге становится меньше. Образно говоря, если вы хотите быть здоровым и энергичным, начните спать с новым партнёром, и имя этого партнера — голод.
Когда вы начинаете практиковать лечебное голодание с оптимизированными периодами, это запускает в вашем организме удивительный механизм из целого ряда биохимических реакций, обеспечивая многогранный anti-age эффект.
Особенность организмов животных, в отличие от растений, в том что для постоянного полного самообновления «точек обновления» должны полноценно срабатывать слаженные механизмы репарации, аутофагии и апоптоза. Только они смогут заменить точки роста, которыми обновляются растения. Известно, что растения в принципе имеют потенцию на неограниченное существование. Например, известна вегетативная роща от одного дерева тополя осиноподобного, возраст которого около 80000 лет. Признаков старения у неё нет. В животном мире такая свобода ограничена рядом правил связанных с институированием онтогенеза. В свою очередь, концепция лимита Хейфлика устаревает, а феномен репликативной остановки клеток имеет другую подоплёку и его можно разблокировать, то есть причины сенесцента клеток всё больше получают опровержения. Это означает, что если нет программы ограничения на уровне клеток, то тогда остаётся реальной стратегия на устранение институированных препон путём приведения в баланс процессов реювенилизации и точек самовосстановления.
В связи с непрерывной динамкой онтогенеза происходит потребность непрерывных перестроек для усиления мощи регулировок одних систем на фоне неизбежного наращивания потребности тормозящих систем ведёт к перегрузке гипоталамуса и делает этот орган самоограничивающимся, а значит настроенным на старение. Гипоталамусу также как и клеткам необходимо давать возможность на полное самовостановление, аналогично тому как это происходит в матрикальной главной почке дерева, которая постоянно перевосстанавливается на новом месте.
Итак, одним из институирующих механизмов старения организма является гипоталамус. Ниже будет приведен обзор доказательных фактов.
Генетические механизмы, через которые аутофагия сдерживает гипоталамическое старение
Аутофагия – это строго контролируемый процесс, который регулирует круговорот клеточных компонентов и поддержание клеточного гомеостаза. С возрастом активность аутофагии снижается, а это вносит свой вклад в различные фенотипы старения и сопровождающие их анизомалии. Можно утверждать, что ослабление потенциала к аутофагии – пролог к клеточному сенесценту в пределах всего организма, но лидером в этих процессах, ведущей скрипкой становится гипоталамус. Он и определяет свои особенности старения у каждого вида животных и их особей. Так, аутофагии способствует повышенная экспрессия NPY в нейронах гипоталамуса. Эта NPY-зависимая стимуляция аутофагии опосредована рецепторами NPY Y1 и Y5 через действие путей PI3K, MAPK/ERK и PKA. Экспрессия NPY в гипоталамусе старых мышей снижается. Заметное накопление p62 в нейронах POMC указывает на то, что нейроны POMC проявляют повышенную чувствительность к возрастному снижению аутофагии, что у старых мышей приводит к увеличению уровней препропротеина POMC в гипоталамусе и снижению a-MSH. Кроме того, у старых мышей обнаруживаются сходные дефекты ожирения и липолиза, как у мышей с нокаутом ATG7, о чем свидетельствуют увеличение массы тела, количества жира и снижение содержания свободных жирных кислот в сыворотке крови и глицерина по сравнению с молодыми мышами. Таким образом, аутофагия нейронов POMC важна для поддержания энергетического баланса за счет генерации a-MSH, а снижение аутофагии в POMC способствует метаболическим осложнениям, часто наблюдаемым при старении.
Гипоталамические корни атеросклероза
Существует убедительная концепция связи образования холестериновых бляшек с возрастным нарушением работы гипоталамуса – руководителя, диспетчерского центра. В силу нарушения обменной и гуморальной регулировки, сигналы макрофагам подаются неточные, в результате чего они объедаются холестерином, поступающим в кровь, превращаются в пенные клетки и обездвиживаются под эпителием сосуда, образуя ту самую холестериновую бляшку.
Гарбузовым Г. А. выдвинута концепция, что механизмы старения организма имеют двустороннюю направленность движения: с верхних этажей регулировок = гипоталамус, как перехода онтостадийных и гормональных перестроек, и с нижнего клеточно-тканного уровня, с которым связан ограниченный клеточный потенциал к репарациям, а на уровне тканей - к регенерациям. Это связано с геномно- эпигеномными перестройками или подстройками под онтостадийные изменения на верхних общеорганизменных уровнях.
На верхнем общеорганизменном уровне это связано с элевационными перестройками в работе гипоталамуса. Изменения в нём связаны как с онтостадийными изменениями, когда «партитура» играет иной гормональный профиль, определяющими онтогенез, так и с банальными гипоксиями гипоталамуса и эпифиза, что является дополнительной причиной снижения их чувствительности к сигналам обратной связи и нарушениями в регулировках.
Гипоксия – недостаток кислорода = кислородное голодание, т.к. в этом случае у тканей и органов наблюдается дисфункция от дефицита кислорода.
Гипоксия гипоталамуса. Она связана с плохим поступлением кислорода из крови.
Итак, в гипоталамусе постепенно происходит снижение притока кислорода. Кроме того, известно что микрокапиллярная сосудистая сеть с возрастом вымирает на 50%, что тоже существенно сказывается на работе мозговых структур, когда страдают как сами крупные сосуды, так и ткани мозга.
В этом случае в гипоталамусе происходят компенсаторные приспособительные процессы, перестройки ограничивающие его возможности как регуляторного централизованного диспетчерского пункта.
Другими словами одним из механизмов этого является хроническая ишемия мозга. Если «копать» глубже в причинах и механизмах патологий гипоталамуса, то можно увидеть «вклад» в это остеохондроза шейного отдела позвоночника.
Гипоталамус навигатор-координатор статуса гормезиса = благополучия
В этом обзоре внимание акцентировано на природу и причины старения гипоталамуса и показано, что оно связано в большей степени не с внешними факторами, а исключительно внутренними процессами саморазвития в нём, неизбежной элевационной динамикой.
Воспаление гипоталамуса одна из причин гипоталамического синдрома. Провоспаление в области гипоталамуса – одна из причин его резистентности, нечувствительности, толерантности его центров регулировок к корректирующим сигналам. Именно первичные проблемы в гипоталамусе являются причиной ускоренного старения организма, а также многочисленной хронической неизлечимой патологии. Следует понимать, что на эти проблемы могут сказываться как экзогенные факторы, вызывающие различную патологию, так и неминуемая эндогенная возрастная динамика в нём. С последними связаны анизомалии = возраст-зависимые заболевания, климакс, старение.
Гормезис реализуется за счет поддержки врождёнными механизмами ВИТАУКТА, то есть на уровне всего организма.
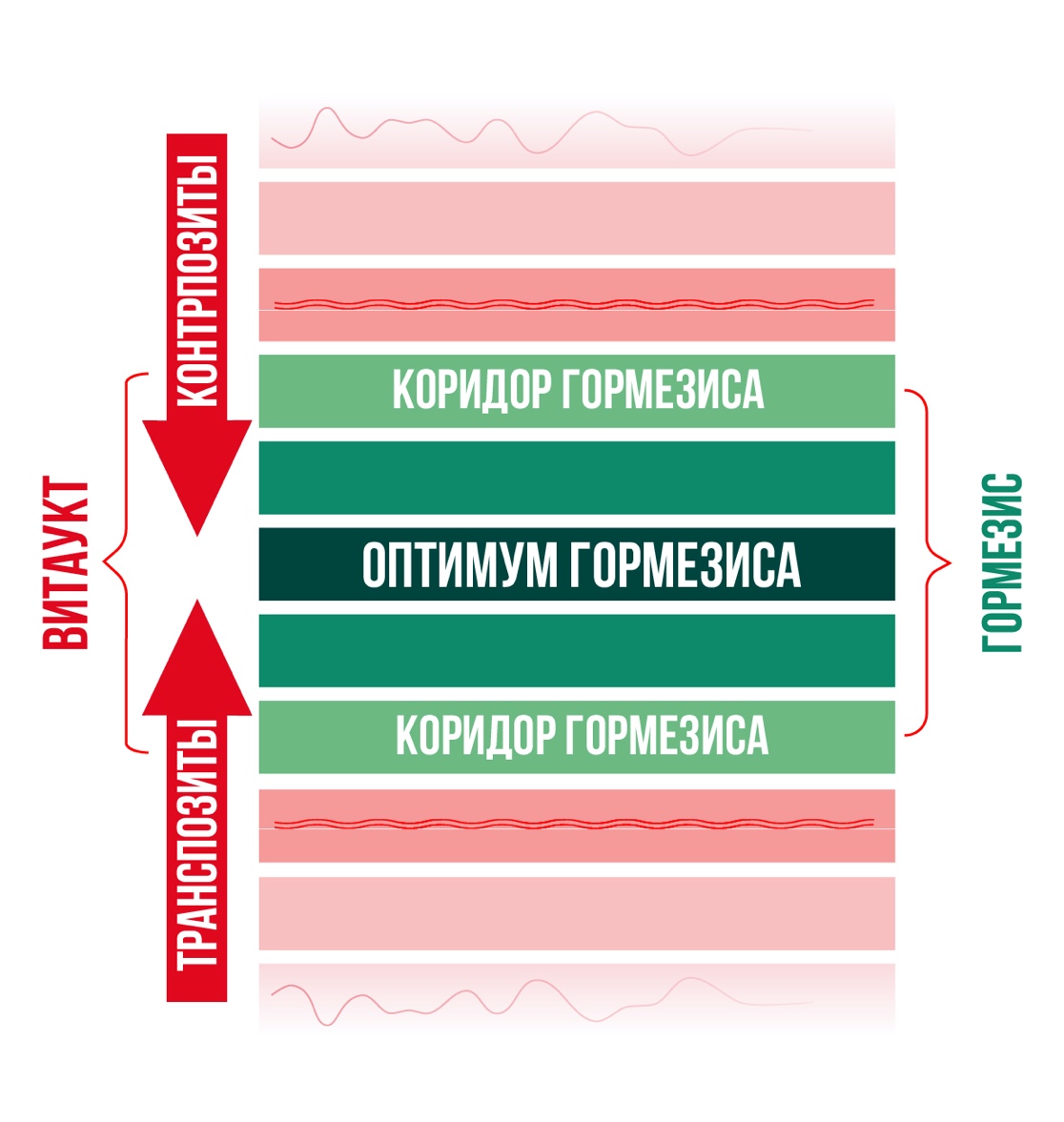
Рис. 4. «Коридор гормезиса», который находится в центре, где работа всех гомеостазов оптимизирована и синхронна, за счет высшей степени проявления возможностей ВИТАУКТА.
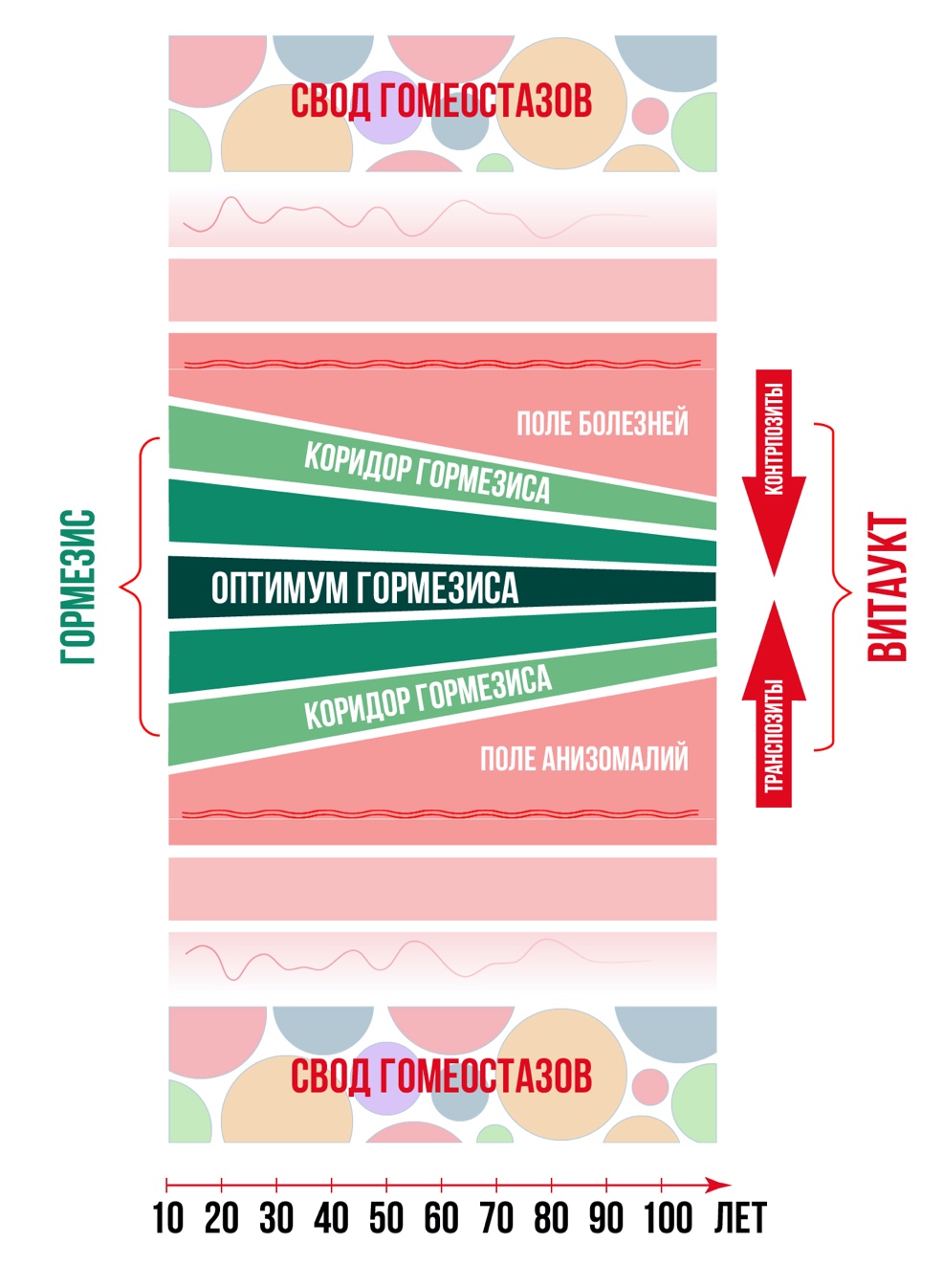
Рис. 5. Показано что «Коридор Гормезиса» у людей с возрастом в онтогенезе сужается, а ВИТАУКТ = Жизненная Сила в виде суммы сил противоположно направленных транспозитных и контрпозитных рычагов гомеостазов уменьшается, «тухнет». Возможности Витаукт противостоять нагрузкам и проявлять адекватную резистентность, чувствительность к регулировочным сигналам в разных системах снижается. За коридором гормезиса открывается супрафизиологический горизонт анизомалий – возраст-зависимых заболеваний, где сил Витаукта недостаточно для противостояния разрушительным процессам.
Напомню, что существуют механизмы регулировок на уровне клеток и тканей, которые обслуживаются механизмами гомеостаза. Гормезис – это режим оптимальной работы всех гомеостазов и систем, всего организма. У гормезиса есть главный инструмент регулировок – это гипоталамус – Генштаб, дирижёр, которому дана синхронизация сводного оркестра и коррекция ему подвластных многочисленных инструментов регулировок. Гипоталамус это, по сути, регулировочный «прибор», который отслеживает всё множество констант, которые нужно поддерживать на заданном уровне, а затем он интегрирует и оптимизирует работу всего этого множества. Таким образом, его следует рассматривать как единую целую структуру, отвечающую за единство всех многочисленных процессов.
Границы возможных регулировочных ответов гипоталамуса заложены в морфогенезе, то есть изначально в нём предусмотрены параметры, на которые он должен реагировать стандартным обратным ответом, не допускающей перекосов в данной системе, сохраняя её постоянство. Таким образом, эта реакция является механизмом, который работает аналогично программе с предусмотренным заранее ответом. Филогенетически заранее предусмотрены регулировочные ответы на то как организм должен отвечать на колебания данных параметров. Таким образом, само наличие гипоталамуса и его реакции является проявлением программ на верхнем уроне регулировок организма. Гипоталамус, это по сути сводный аналитический штаб, диспетчерско-контрольный центр многочисленных регуляторных пунктов, осуществляет функции гормезиатора-интегратора = гармонизатора-синтезатора. Точно также как на нижнем уровне работают механизмы гомеостата, так на верхнем работает – гормезиостат. Таким образом, баланс всех общеорганизменных процессов регулируется сверху на уровне гипоталамуса, включая подконтрольные ему системы: нейроэндокринные и иммунную. Над всеми ими стоят центры регулировки и сохранения гормезиса – гармонии. Это проявляется в оптимальном соотношении работы всех механизмов Витаукта, когда организм находится в идеальном состоянии.
Аналогия регулировочной системы гипоталамуса с системой ГЛОНАСС
Для наглядности посмотрим на гипоталамус в сравнении с системой ГЛОНАСС - глобальная навигационная спутниковая система. Это спутниковая система навигации, через космос управляющая всей транспортной системой и её потоками. Нечто подобное осуществляет и система гироскопов в самолётах, но в клетках эту функцию контроля и навигации всех других интегральных систем осуществляют белки-интегрины. Аналогичную систему регулировок с помощью интегринов на уровне клетки Гарбузовым предложено называть ГЛОНАСК - глобальная навигационная система клетки, а на уровне организма ГЛОНАСО - глобальная навигационная система организма. Таким образом, гипоталамус можно полноправно назвать молекулярным ГЛОНАСО. Цель их держать постоянство, правильный курс всей системы.
На клеточном уровне работают свои механизмы регуляторы – гомеостаза, а на общеорганизменном уровне работают другие механизмы регулировок - гормезиса. Регулировка идёт с двух направлений: сверху и снизу, то есть как со стороны ГЛОНАСК, так и ГЛОНАСО. Другими словами ГЛОНАСО можно обозначить как гормезис-стабилизационная-система организма = ГОРМЕСТАБСО. Таким образом, автор предложил во избежание путаницы и двусмысленности вести новый термин: ГОРМЕСТАБСО.
Аналогично на уровне клетки это будет ГОМЕСТАБСК – гомеостаз-стабилизационная система клетки.
Итак, у организма имеется два уровня механизмов сохранения постоянства: ГОРМЕСТАБСО и ГОМЕСТАБСК. Сочетание этих систем создаёт единую систему обеспечения здоровья организма – ВИТАУКТ. Витаукт состоит из равновесия множества механизмов, у каждого из которых имеются противоположно направленные силы: транспозиты и контрпозиты, цель которых вернуть любую систему к точке константы присущую для каждого гомеостаза.
Верхние и нижние этажи регулировок онтогенеза, болезней старости и старения
В этом аспекте большое значение для практики и теории имеет получить ответ на вопрос какие уровни первичны и имеют первостепенное значение в понимании проблемы механизмов онтогенеза и старения организма, а также возраст-зависимых заболеваний = анизомалий, откуда идут истинные начала, корни проблемы - сверху или снизу.
На уровне гомеостазов проявляются процессы сенесцента клеток (клеточного старения), а на уровне гормезиса – сенилит – старения исходящего с верхних уровней регулировок организма, и обусловленного своими возможностями регулировок, своими программами и механизмами. Вопрос: кто кого обуславливает, или каждый из них вносит свою лепту в процесс старения и сопровождающих его анизомалий? Значит гомеостазы и гормезис не постоянны и в онтогенезе смещают свои параметры, причём выходят за рамки оптимума. Динамический гормезиорез и гомеорез, термины которые определяют это понятие. Подробности в книге Гарбузова Г.А.: «БИОЛОГИЯ МОЛОДОСТИ».
Факты указывают на то, что старение это двойственный поток процессов: сверху и снизу. В первом случае с верхней стороны-уровня нисходят свои институированные установки-предписания регулировок, в конечном итоге которые ведут к старению. Эти установки-предписания являются по сути программативными для подчинённых структур. Институированное управление означает, что в организме имеются центральные органы, как дирижёр управляющие всей разноголосицей многочисленных систем и приводящих их в симфонию единого оркестра. Все органы и гомеостазы работают не сами по себе, а взаимоувязаны между собой и всеми так различными механизмами координации, что играют когерентно в такт строго под одну мелодию, создавая полную гармонию. Ни одна система не может сыграть за пределами общей мелодии.
С другой стороны происходят восходящие процессы, ограничивающие репликационные возможности клеток, а значит неограниченное самовосстановление тканей и существование всего организма. Это другой этаж ограничивающий возможности к неограниченному самовосстановлению.
Прямым доказательством того, что старение обуславливает верхний гипоталамический этаж регулировок является эксперимент когда молодую и старую мышь сшивали так, что у них начала работать единая кровеносная система. Это так называемый парабиоз.
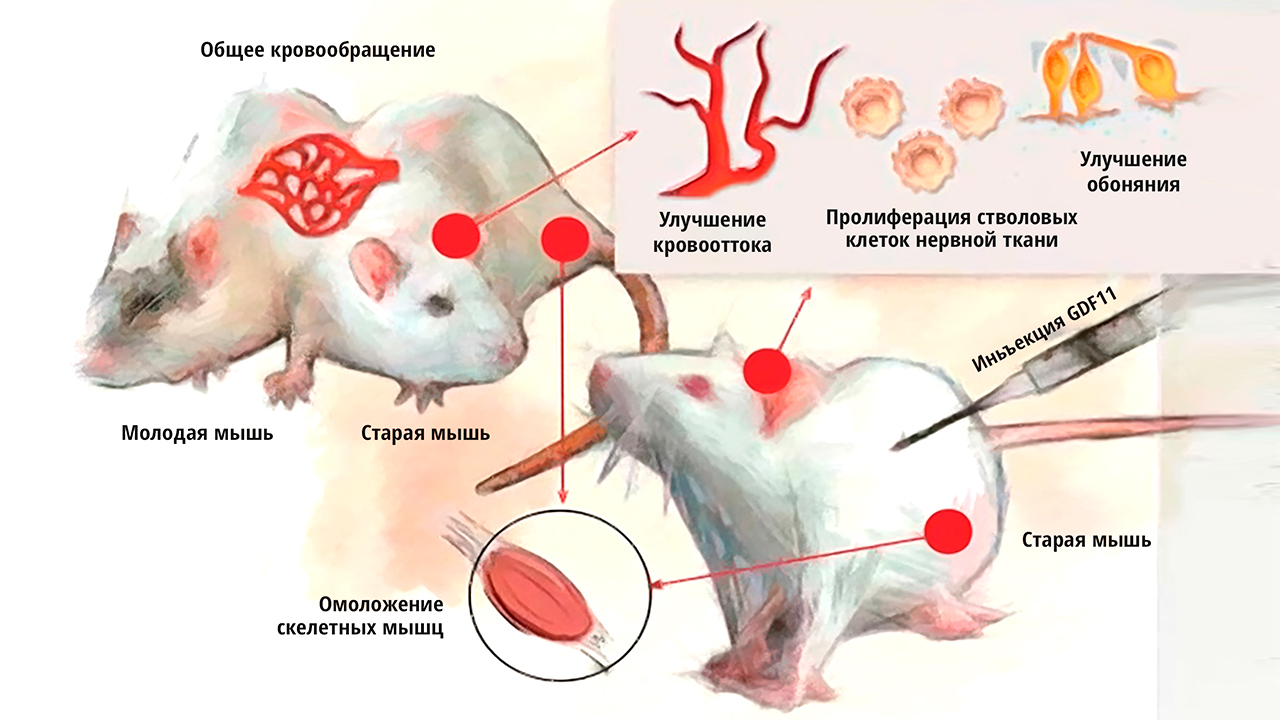
Рис. 6. Молодая и старая мыши, у которых сшита воедино кровеносная система, и таким образом выявлен эффект гетерохронического парабиоза, когда за счёт молодой крысы удалось частично омолодить старую, но при этом состаривалась и молодая особь.
При этом старая мышь приобретала множество признаков омоложения её организма, а молодая мышь при этом ускоренно старела. Следовательно, в крови у старой мыши появился некие вещества-регуляторы, которые старят весь организм, а у молодой мыши есть вещества, которые омолаживают старую. Это означает, что если мы сможем выделить это вещество, то получим ключ по омоложению всего организма.
Подтверждением этому являются данные о том, что между мозгом и телом есть какие-то вещества ускоряющие или сдерживающие старение. Было найдено что в гипоталамусе мышей есть клетки, которые влияют на скорость старения.
Выяснилось, что в процессе старения активно задействованы нервные стволовые клетки (НСК). Когда у некоторых мышей среднего возраста их убрали, животные очень быстро постарели — значительно быстрее, чем в обычной ситуации. Потом престарелым мышам из другой группы ввели НСК новорожденных мышей. Через некоторое время все «пенсионеры» стали более подвижными, выносливыми, у них явно улучшились когнитивные способности.
Обнаружен белковый элемент, без которого никакого омолаживания нет — это белок TIMP2. Он выполняет функцию блокатора металлопротеиназ. Если его убрать из плазмы, то никакого эффекта нет.
Кроме того, у стареющих мышей проблемой являлось уменьшение числа стволовых клеток как во всём организме, так и в самом гипоталамусе. Как результат вместо мезенхимных клеток стали преобладать второстепенные клетки помощники – прогениторные. Автор предложил это обозначить как эффект соматического доминирования. Гипоталамус стареет наравне со всеми остальными структурами, благодаря первичным клеточным процессам сенесцента, но при этом гипоталамус является системой-лидером, от функциональности которой зависят ткани-мишени. Поэтому, старея сам, гипоталамус еще и ускоряет дополнительно старение других тканей. Поэтому гипоталамус имеет двойную роль в процессах сенилита: как институционную, так и сенесцентную.
Итак, кроме верхнего институционного уровня старения имеется и всеобщий клеточный уровень – сенесцентный, который несёт свою лепту в общий процесс старения. Очевидно, он связан с найденным клеточным гормоном MOTS, который вырабатывается в митохондриях, а не в ядре клетки. Мыши которые получали инъекции гормона, чувствовали себя гораздо лучше всех остальных. При этом старые мыши, получившие серию таких инъекций, стали ходить по беговой дорожке вдвое быстрее. У них даже получилось обогнать гораздо более молодых животных, которые не получали никаких инъекций.
Таким образом, приведены данные подтверждают, что процесс старения зарождается как с нижнего, так и верхнего этажей регулировок организма.
Что такое геронтос или взаимоотношения процессов старения на верхнем гормональном уровне организма (сенилит) с процессами старения на уровне клеток (сенесцент)
Итак, мы показали, что процессы старения организма (сенилит) в целом регулируются на уровне гипоталамуса, а на уровне клеток старение (сенесцент) за счет транскрипции и активизации или угнетения, блокирования теломеразы. Теломераза фермент реализующий возможности восстановления теломер. Но теломераза тоже находится под контролем и зависит от множества факторов. У неё есть свои рецепторы регулировок, но в области эпигенома. И эти рецепторы настроены не только сами на себя, контроль внутренних процессов, но и на внешние сигналы, определяющие статус гормезиса. Между ними должны быть сквозные взаимоотношения, пронизывание; между гормезисом и гомеостазами. Если точнее, то имеется в виду не все гомеостазы, а те которые обеспечивают, контролируют сенесцент. Длина теломер и скорость их укорочения это не застывший параметр, а регулируемый и нельзя говорить, что укорочение теломер это односторонний процесс, который не поддается обратному движению. Напомню, что в онкоклетках происходит быстрая перестройка-подстройка этого процесса под новые условия: онкоклетка попросту полностью отключает программу на сенесцент, включая программу транскрипции теломеразы. Итак, длина теломер это регулируемый параметр, где всё зависит от того на какую величину этот параметр настроен, запрограммирован. В растительном мире этой блокировки числа репликаций клеток вообще нет, потенциально они могут пролиферировать бесконечно, нет сенесцента. На продолжительность жизни растений клеточный уровень не влияет вообще, все определяется с верхних уровней апикальных меристем. Это так называемый принцип институционного регулирования организма, то есть гормонами, нейромедиаторами. В мире животных появляется дополнительный уровень регулировок старения – клеточный, через лимит Хейфлика. Также у животных появляется принцип коммитации, то есть с обязательством, заложенной внутри клетки программой производить только один тип клеток, включены только на одну потенцию, например, унипотентные клетки или выполняют только один функционал или морфогенную единицу. Подробности в книге Гарбузова Г.А.[2].
Лимит Хейфлика связан не с тем, что клетка, изнашивается, исчерпывает какие-то свои ресурсы, а с тем что она настроена, запрограммирована так. У каждого вида животных имеется свой параметр числа репликаций, свой лимит Хейфлика. Но всегда выключена функция активирования теломеразы. Для этого есть специальные свои механизмы и программы. Все параметры заранее предопределены природой.
С возрастом длина теломер укорачивается. У человека длина теломер варьирует от 15 тысяч нуклеотидных пар (т.н.п.) при рождении до 5 т.н.п. при хронических заболеваниях. Длина теломер максимальна у 18-месячных детей, а затем она быстро снижается до 12 т.н.п. к пятилетнему возрасту. После этого скорость укорачивания несколько снижается. Это усиливает сенесцентность клеток. Темпы старения клеток регулируются изнутри на уровне механизма хромосом. Вместе с нарастанием сенесцента снижается количество стволовых клеток и уровень их потентности; они становятся более резистентными, нечувствительными, слабо функциональными.
Но эта скорость сенесцента, связанная с длиной теломер, не стабильный параметр и отчасти варьирует у разных людей. На эту скорость значительно влияют стрессы, то есть факторы извне, а также выявлены многочисленные факторы, которые могут на неё повлиять. Число репликаций в культуре ткани иногда удаётся поднять с 50 до 262. Это говорит о том, что у клетки есть рецепторы, которые чувствуют внешнюю среду и отвечают на неё. В свою очередь у гипоталамуса имеется группа центров-ядер, которые отвечают за горметическую стабильность, а на них рецепторы ощущающие динамику на гомеостазах отвечающих за статус сенесцента в них. Сенесцент и сенилит – взаимозависимые процессы, обуславливающие друг друга, между ними должны быть сквозные взаимоотношения. По сути, это главная ось регулировки онтогенеза, возрастных динамических перестроек.
Клеточное старение (сенесцент) и общеорганизменное старение (сенилит) демонстрирует эффект отрицательной плейотропии, суть которого состоит в том, что то что хорошо для молодого организма, может стать плохим для старого. Самый яркий пример — процессы воспаления. Процессы провоспаления инициируются как с верхних этажей регулировок, в частности со стороны иммунитета, так и с клеточного уровня.
Гормонозависимые опухоли как проявление сенилита и сенесцента
Показательным примером возрастных динамических перестроек и взаимосвязи сенесцента и сенилита в качестве главной оси регулировок онтогенеза могут быть гормонозависимые доброкачественные опухоли (мастопатия, миома, аденома) и злокачественные опухоли (рак груди, простаты…). Зачатки их зарождаются в условиях хронического провоспаления, одним из возбуждающих факторов которого является изменение с возрастом гормонального фона. Особо это выражено на примере снижения мелатонина до 10 раз на последних стадиях онтогенеза адальтус, матура, сенилит. В данном случае тип старения человека имеет частичное сходство со старением рыбы горбуша, у которой гипергипоталамическое доминирование ведёт к неизбежному гипергонадизму, гипернарастанию половых гормонов с исходом в феноптоз, то есть самоуничтожения организма. По сути этот гипергонадный тип старения аналогичен монокарпическому старению и феноптозу у агав.
Гипергонадизм причина гормонозависимых опухолей у человека.
Эти виды опухолей являются одним из проявлений анизомалий, то есть факторов для феноптоза. Эта причина всегда двойственна: неизбежная гиперактивация гипоталамуса на фоне ослабления корректирующего, смягчающего действия со стороны эпифиза и недостатка мелатонина (элевационные процессы в нём) и переход на форсированные режимы нагрузки, что ведёт к гиперфункциям на железах-мишенях.
Возрастное угасание мелатонинового рычага для коррекции гипоталамуса как фактор сенилита.
В образовании анизомалийной опухоли важна не просто гиперфункция железы-мишени, вырабатывающая избыточное количество половых гормонов, но и недостаточное сопровождение мелатонинового фона; то есть нарушены не просто количественные показатели гормонов, а именно их соотношения с мелатонином, работающего как противовес. Есть оптимальные величины показателей их соотношения, которые соблюдены в молодости. Но в ряде случаев их пределы и соотношения выходят за рамки оптимума, такая система выходит как за пределы гормезиса, так и за пределы клеточных гомеостазов. Зрелость гипоталамуса, повышение его чувствительности происходит при снижении корректирующего, тормозящего действия эпифиза.
Компенсационные возможности мелатонина для сдерживания анизомалийных опухолей.
Ряд данных применения мелатонина для повышения эффективности терапии раковой опухоли показывают, что для их сдерживания нужны не обычные дозы 1,5 – 5,0 мг, рекомендуемые для улучшения сна, а на много больше – 20 – 40 мг. Очевидно такие высокие уровни мелатонина специфичны для ранней юности, когда происходит ювенильное доминирование эпифиза над гипоталамусом. Кроме того, в зрелом возрасте на фоне развившегося гипергонадизма обычные дозы мелатонина, которые приемлемы в детстве, уже не достаточны. Гормонозависимых опухолей в этом возрасте не бывает. Раковые клетки как и сенесцентные обладают высокой степенью резистентности как к их подавлению, так и к мелатонину. Это связано с нарушением настройки рецепторной системы на мембранах и хроматине (сенсорные нарушения чувствительности к сигналингу из-за эпигенетических перестроек на хроматине). Методы гормональной кастрации применяемые при гормонозависимых опухолях, к сожалению, это соотношение гормонов с мелатонином не учитывает и поэтому получает намного ниже результаты от желаемых.
Чем слабее эпифиз, тем сильнее гиперфункция гипоталамуса, выше его гиперэлевационное состояние и наоборот, чем сильнее гипоталамус – тем слабее эпифиз. Удаление эпифиза в детстве ведёт к преждевременному созреванию. Причём вредна не сама активность, а его гиперактивность, перезрелость. Эпифиз ведёт гипоталамус, «протаскивает» его по всем ступенькам онтогенеза от ювенала до адальтуса и матуры. Если точнее, то «протаскивает» до стадии юности, а затем происходит перенастройка и гипоталамус подавляет эпифиз. Любая гиперфункция - это всегда выход за пределы оптимума, за пределы коридора гормезиса, функционально расточительна и всегда переходит в другую крайность - в дегенеративный исход и гипофункцию. Причиной катастрофической перезрелости и неизбежности смертного исхода у рыбы горбуши является даже не сенилит, а гиперадальтус – избыток гормонов. Движение до матуры (климакса) даже не доходит. Происходит перенапряжение ряда желез, когда они работают за пределами констант, что неминуемо запускает механизмы самовоспаления. Это является первым шагом к открытию шлюзов и прологом для маршрута к их онкологизации. У человека неизбежной причиной мортуса (смертности) являются:
Такие раковые опухоли как рак матки, рак груди, рак простаты являются детищем гормональных перестроек, гипергонадизма, то есть корни их исходят из внутренней дисгармонии, которые усиливают дополнительно гормоны стресса и ослабление мелатонинового буфера. Например, опухоль простаты – аденома увеличивает вероятность своего проявления с каждым следующим 10-летием. Так вероятность аденомы или гиперплазии простаты в 50-летнем возрасте 30-50%, а в 60 лет – 50-60%, 70 лет – 70-80%, а в 90-100 лет достигает 100%. Таким образом, аденому не верно называть болезнью (связана с внешними факторами), это типичная анизомалия (неизбежные проявления на основе патологопредраспологающих процессов в разных системах организма, связанные с внутренними перестройками). С возрастом с нарастанием проанизомалийных перестроек = антигормезисных увеличивается предрасположенность к онкологизации ткани. Одновременно повышается степень сенесцентирования на клеточном уровне. В гипоталамусе сенесцент клеток является причиной его нечувствительности к тормозящим сигналам обратной связи, что в свою очередь ведёт к гипергипоталамическому доминированию. Любая преданизомалийная перестройка связана с митохондриальными дисфункциями: сначала гипер-, а затем гипо-. Это основа сенесцента, который в свою очередь может повести по двум направлениям:
- онкологическая настроенность,
- сенесцентное дегенерирование ткани.
Это две крайности спектра проявлений одного и того же процесса. Всё зависит от того какое крыло будет превалировать – то по тому руслу и пойдет процесс.
Противовесом, контрпозитной силой, очевидно, в этом случае выступают механизмы старения клеток, сенесцента. Это режим перевода данных клеток в состояние более мягкого провоспаления, чем при онкологии. Это компенсаторный механизм сглаживания остроты данной анизомалии, позволяет организму получить еще добавочное время в виде нескольких лет или десятилетий.
Сходство раковых и сенесцентных клеток.
В любом случае и онкологические клетки, и сенесцентные имеют сходство в том, что работают в гликолизе и у обоих процессов проявляется митохондриальная дисфункция, недостаточность.
Кроме того, не верно утверждать, что сенесцентирование это механизм защиты от онкологизационного пути развития. Сенесцент – это всего лишь следствие преданизомалийных перестроек и связанных с ними митохондриальных дисфункций, которые происходят по принципу компенсационных гиперфункций, а затем их гипофункций и последующего сенесцента. Возрастная тенденция предрасположенности к онкологизации - это всего лишь один из путей, вариантов проявления анизомалийных перестроек, связанных с выходом за границы коридора гормезиса. Сенесцент – это следствие и другой вариант проявления преданизомалийных перестроек. Это подразумевает, что каждая онтостадия развития организма готовит свои условия как для сенесцентирования клеток, так и для онкологического перерождения. А не наоборот, когда утверждается что первично происходит сенесцент, и он обеспечивает анизомалийные перестройки. Из этого следует, что сенесцент должен идти шлейфом за гипоталамическими перестройками.
Доказательство того, что сенесцент это следствие онтостадийного продвижения и сенилита
Таковым является то, что у долгоживущих типов животных и человека процессы сенесцента, анизомалийные перестройки и анизомалические патологии проявляются намного в более старшем возрасте – после 40-60 лет, тогда как у короткоживущих животных (мыши) тот же самый спектр анизомалийных болезней, но сенилит проявляются после 2 лет. Так анизомалийные патологии как диабет, атеросклероз, онкология... у мышей начинают появляться в возрасте 1-1,5 года, тогда как у человека в этом возрасте нет и зачатков этих болезней. Причиной является выход в поле анизомалий, то есть за пределы коридора гормезиса. Это означает, что сенилит начинается на общеорганизменном уровне, к которому присоединяется и клеточный уровень – сенесцент. Следовательно, не клеточный уровень сенесцента определяет длительность жизни, а особенность реализации онтостадийных становлений организма. Именно особенности гипоталамуса, а также возможности мозговых тканей обеспечить трофику тканей мишеней, являются драйвером онтогенеза и определяют способность к долгожительству или короткожительству животного. Верхние уровни регулировок определяют клеточный сенесцент, а не наоборот.
Сенесцент клеток это программа, которая возникает и обеспечивает существование клеток в условиях частично находящихся за пределами гомеостаза, чему в свою очередь способствуют условия выхода организма за пределы гормезиса. Имеется в виду переходы на последние этапы онтогенеза. Данные по лимиту Хейфлика, которые противоречат предлагаемой нами концепции, являются всего лишь артефактом, когда результаты эксперимента были не правильно трактованными. Клетки животных в культуре ткани, которым обеспечен гомеостаз, тем не менее, находятся за пределами гормезиса. Как результат – они частично находятся в поле анизомалий. Чем дальше продвинуты этапы онтогенеза, тем глубже клетки окунаются в поле анизомалий. В отличие от клеток растений, клеткам животных нужны условия не только гомеостаза, но и гормезиса. Эта особенность обусловлена тем, что регулировка ряда онтостадийных процессов у них вынесена с общеорганизменного уровня (институировано, то есть с помощью гормонов) на клеточный уровень (коммитировано). Такие клетки заведомо самоограничены на предмет повышения потентности. Это в свою очередь снижает диапазон маневрирования степенями потентности стволовых клеток. Только высокопотентным клеткам дана возможность на полную репарацию, в том числе и митохондрий, и эпигенома. Клетки с низкопотентными способностями несут в себе весь груз онтостадийных перестроек, а значит лимитированы на сенесцентность. Такие клетки будут зависимыми от некоторых внешних факторов, которых не хватает в культуре ткани.
Да, сенесцент исходит из нижнего клеточного уровня, но он не изначален и не ведущий механизм старения, а всего лишь коммитированное продолжение общих процессов.
Тем не менее, сенесцент проявляется на нижнем клеточном уровне, а сенилит – общее старение нисходит с верху. Это два процесса происходящие на разных уровнях регулировок, но это две стороны одного общего процесса. У сенесцента есть свои программы и платформа, но они запускаются на рельсах онтостадийных становлений. Сенилит через сенесцент реализует свои лимиты.
Сенилит является институционным процессом, а сенесцент – коммитированным. Таким образом стадия онтогенеза – старение регулируется с двух сторон, и это более плавная корректировка чем при одностороннем регулировании сверху.
Каждая генетическая программа коммитирования основана тоже на принципах доминирования какой-либо гено-программы, которая и переподчиняет себе всё остальное множество программ. Это доминирование регулируется уже не внешними, а внутри клеточными гормонами. Также как и при апикальном доминировании у растений, когда его можно снять с помощью гормона цитокинина, сенесцентное доминирование тоже можно снять, отменить с помощью аналогичного гормона у животных. Цитокинин способен отменить как апикальное доминирование, так и онтофизисное состояние усыхающих старых ветвей. Таким образом, такой универсальный гормон как цитокинин смог бы отменять как институционные, так и коммитированные настройки. Это позволяет управлять как процессами регенерации, на уровне тканей и органов, так и клеточной репарацией.
В культуре ткани клетки животного типа имеют направленное ускорение на сенесцент, что обусловлено коммитационно.
Драйвером элевационных процессов в гипоталамусе является потребность в непрерывной динамике, его развитии, созревании, а также в связи с необходимостью компенсировать жесткое торможение роста организма. Современный животный мир приобрел особые свойства гомойотопии — ограниченность роста (от слова «гомойо» — способность всех особей вида или органов в них достигать жёстко заданных величин, например размеров, а «топо» — от слова «топология» — размеры, пространство). Гипоталамус с возрастом вынужден непрерывно наращивать механизмы торможения роста за счет усиления генеративных функций. То есть, на каждом этапе развития происходит диспропорция гормонального фона от предыдущего, изменения идут не количественные, а качественные. Эра эволюции, когда животные существовали по принципу пойкилотопии* (неограниченного роста и гигантских размеров), канула в лета. Чем сильнее притормаживается рост, тем сильнее проявляется зрелость организма, такое наращивание зрелости переводит организм с этапа юности на этап зрелости = адальтуса, в дальнейшей динамике адальтус ведёт к перезрелости, то есть переходу на этап матуры, а он является прологом к сенилиту = старости.
Каждая специализированная ткань имеет свой предел гомойотопии. Этому ограничению не подвержены только стволовые клетки крови, лимфы, половые… Эта гомойотопия связана с клеточным коммитированием, то есть своеобразной «подстраховки» регулировки снизу процессов развития. Коммитирование и сенесцент в свою очередь являются следствием эпигенеза.
Доказательство того, что старение идёт сверху, а не снизу.
Таковым является эксперимент, когда проводили пересадку яичника от старой крысы молодой и при этом происходило восстановление циклической работы яичника. Клеточный лимит Хейфлика здесь оказался ни причём. Следовательно, лимит клеток старого яичника не был исчерпан, просто работа яичника была заглушена на гормональном уровне. В свою очередь пересадка яичника от молодой крысы в старую к результату не приводили – цикл яичника не восстанавливался. «Молодые» клетки переподчинились регулировочному циклу старого организма. Вывод, что в геронтосе* (единство сенесцента ⇆ сенилита) по оси гипоталамус-яичники первопричиной геронтоса является не сенесцент, а сенилит.
Старение яичников вторично, а не является следствием сенесцента. Старение яичника это проявление климакса, а значит климакс тоже вторичен и является следствием сенилита. Сенилит это производная перестроек гипоталамуса. А допустим если там нет перестроек, то это означает, что нет и сенилита.
В медиа прессе описан случай с нестареющей Розой Фарони, которая в 97 лет выглядела на 30-летнюю. Если присмотреться к её случаю, то можно заметить отсутствие климакса, то есть сохранился менструальный цикл. А это означает, что гормональный женский фон у неё сохранился. Возрастные элевационные перестройки в её гипоталамусе ни качественные, ни количественные не произошли. Значит принципиальных препятствий не элевационирования гипоталамуса нет? Он может длительно самообновляться (за счёт стволовых клеток в точках самовозобновления) и неограниченно поддерживать свой статус молодости или зрелости? Какой-то механизм мешает такой самоподдержке. Не клеточный ли это сенесцент? Или этим механизмом является увядание эпифиза? Эпифиз противофаза гипоталамусу? Он удерживает юность и отодвигает зрелость? Процесс идёт на качелях эпифиз ⇆ гипоталамус? Или это потребность противодействовать плейотопии путём наращивания степени генеративной зрелости, которая подавляет рост?
Если мы сможем ответить на этот вопрос правильно и получить ответ как добиться стабильности в гипоталамусе, не идти по пути непрерывной элевации, то тем самым сможем остановить сенилит. По какой-то причине, «поломке» в гипоталамусе отключился возрастной механизм элевации и он остался работать на прежнем уровне. Тем не менее, это нарушение «неминуемой» динамики гипоталамуса не повлияло никаким образом на возникновение проблем во всех остальных системах организма. Они благополучно оставались молодыми даже в мафусаиловом возрасте. Значит климакс не неизбежен? Произошла отмена элевационных механизмов, а это означает, что эти механизмы не являются эволюционно неизбежными. Эволюции, очевидно, более важна быстрая смена поколений.
Возрастное нарастание степени резистентности гипоталамуса.
В «норме» с возрастом гипоталамус становится резистентным, нечувствительным к тормозным сигналам обратной связи, что растормаживает его и побуждает к усилению функций и это является проявлением его элевационных перестроек. Эта резистентность связана с клеточной рецепторной нечувствительностью. Гипоталамическая ось регулировок выходит из состояния гипоталамического доминирования и переходит на принцип гипергипоталамического доминирования и тем самым задаёт ограничительный пределы. За гипердоминированием (матурой) происходит спад, истощение, сенелирование. Снимите это гипердоминирование (чрезмерность, зашкаливание за гормезис) и онтогенетические правила поменяются. Случай нестареющей женщины аналогичен с удалением генеративного цветочного побега у агавы, описанный нами ниже, что стало причиной отмены явления гиперапикального доминирования, а это стало причиной того, что данное растение-монокарпик изменило свой онтогенез и жизненный цикл её перешел с 8-летнего на 100-летний.
Эпифизарное регулирование как степени зрелости гипоталамуса, так и его нечувствительности в последующем.
До 12 лет у человека эпифиз сдерживает, ограничивает гипергипоталамическое доминирование; затем эпифиз вырождается и проявляется «вспышка» генеративной гиперактивности гипоталамуса, что в конечном итоге ведёт к его адальтусу (зрелости), затем к матуре (гиперзрелости, климаксу), сенилиту… мортусу. Как видим двух-стадийную динамику: резкого всплеска, подъёма, которая затем переходит в обратную фазу – снижение функциональных возможностей и дегенеративный процесс. Динамика идёт по экспоненте: на графике это парабола, вверх – вниз. Внешне у человека это проявляется в виде усыхания его трофики, увядания, дряхлости, старческой немощи и множества анизомалий*. Возможно, эпифиз в описанном случае продлённой молодости с возрастом не отключил свою работу (эпифизарное доминирование?) и тем самым продолжал сдерживать функциональное перезревание гипоталамуса = запредельное гипердоминирование. Очевидно, с возрастом по ряду причин лидерство эпифиза ослабевает, он становится зависимым к подавляющим его факторам. Но при этом усиливается, растормаживается роль гипоталамуса. Тогда следует ответить на вопрос: что означает и почему эпифиз вырождается после 12 лет? Его подавляет становление этапа пубертатного гормонального созревания, новый гормональный фон.
Однозначно можно утверждать, что до 12 лет организм человека существует на принципах эпифизарного доминирования. Затем осуществляется онтостадийный переход на пубертат* и на гипоталамическое доминирование. Пубертат тоже делится на несколько подэтапов и длится 3-4 года. На первом этапе идёт оволоснение вокруг мошонки и подмышками, но без оволоснения выше по животу, только через несколько лет начинает расти борода и волосы груди.
Гипоталамо-эпифизарный паритет.
Это означает, что первый подэтап пубертата тоже еще находится под контролем, а точнее под значительным влияние эпифиза, можно сказать эпифиз и гипоталамус находятся еще в балансе и выработка гормона эпифиза продолжается еще до 16-20 лет. Это и обеспечивает особенности молодости этого возраста, наилучшее качество трофики всех тканей, все ткани находятся под контролем как гипоталамуса, так и эпифиза. Это так называемый гипоталамо-эпифизарный паритет, который обеспечивает лучшее качество жизни с минимумом анизомалийных проявлений и максимумом Жизненной Силы и Витаукта. Если бы мы умели продлевать и сохранять этап эпифизарного доминирования, то могли бы значительно продлить свой ювенальный этап, а если бы удалось поддерживать этап гипоталамо-эпифизарного паритета, то это позволило бы сохранить молодость в зрелом генеративном возрасте. Сохранение же состояния первого этапа пубертата (незаконченный пубертат) и продление его на многие годы вперёд будет говорить о проявлении неотении*.
Возможности мелатонина в подавлении гипергипоталамического синдрома.
Тогда давайте посмотрим на проблему с другой стороны: а что если поставлять в организм гормон эпифиза мелатонин в больших количествах после наступления пубертата? В первую очередь обращает на себя внимание его возможность регулировать нейроэндокринные функции. Избыточная секреция мелатонина клетками пинеальной железы приводит к гипогонадизму. Известно, что мелатонин у женщин благодаря отсрочке в наступлении менопаузы продлевает молодость. Это связано со способностью мелатонина восстанавливать цикличность выработки половых гормонов и поддерживать их уровень на показателях, характерных для молодого возраста. У мужчин отодвигается старение благодаря продлению активного сперматогенеза, поддержанию уровня половых гормонов, в том числе тестостерона, и защите от импотенции.
Даже инволютирующий гипоталамус на этапе климакса можно вернуть в исходное состояние.
Также хорошо известен феномен снижения чувствительности гипоталамуса ко многим сигнальным факторам, что происходит не на этапе его элевации, а на этапе – инволюции. Толерантность, нечувствительность, резистентность проявляется к огромному числу активирующих его факторов, это так называемое старческое усыхание, его увядание или климакс. При этом было показано, что пептидный экстракт эпифиза, восстанавливает у старых самок крыс регулярные эстральные циклы и чувствительность их гипоталамических половых центров к эстрогенам - механизм, которому придается ведущая роль в возрастном выключении репродуктивной функции. Климакс был отодвинут. Важно отметить, что отодвижение климакса показывает возможность влияния гормона эпфиза даже в далеко продвинутых этапах онтогенеза и старости. Считалось, что можно продлить молодость, но нельзя вернуть молодость из старости. Следовательно, гипоталамус увядает, инволютирует не сам по себе, а в связи с отсутствием соответствующей омолаживающей поддержки со стороны эпифиза. Но при этом на фазе своего опережающего роста он сам же своей гиперфункцией подавляет эпифиз, что и открывает дорогу к старению гипоталамуса; если уже гипоталамус вырвался из подчинения эпифиза, то он сам начнёт его угнетать. До поры до времени эпифиз в молодости сдерживает гипоталамус, но затем гипоталамус начнёт сдерживать эпифиз, начинается их антагонизм. Это означает, что задачей предстоит решить проблему не выпускать гипоталамус из-под контроля эпифиза. Но это возможно только на фазе ювенильности организма – расцвета его юности: 12 -16 лет.
Важно подметить, что отмена климакса у старых животных была сопряжена с отодвижением системного сенилита, и что это не было следствием сенесцента клеток. До сих пор многие авторы придают первостепенную значимость сенесценту в геронтосе*.
Физиологическое действие мелатонина.
Мелатонин увеличивает концентрацию гамма-аминомасляной кислоты (ГАМК) и серотонина в среднем мозге и гипоталамусе, изменяет активность пиридоксалькиназы, участвующей в синтезе ГАМК, дофамина и серотонина. Регулирует цикл сон-бодрствование, суточные изменения локомоторной активности и температуры тела, положительно влияет на интеллектуально-мнестические функции мозга, на эмоционально-личностную сферу. Способствует организации биологического ритма и нормализации ночного сна. Улучшает качество сна, ускоряет засыпание.
Мелатонин - это целая группа веществ, обуславливающих плюрипотентное тормозящее воздействие эпифиза на самые различные железы внутренней секреции: половые железы, щитовидную железу, секрецию гонадо- и соматотропного гормонов гипофиза. С другой стороны секреторная активность эпифиза варьирует в зависимости от степени освещенности. При этом ограничение освещенности, а значит увеличение мелатонина в условиях севера связывают проявлением депрессии, которая в свою очередь связана с ГАМК-эргической системой, дофамином и серотонином.
С другой стороны важно понимать как соотносятся данные результаты этого эксперимента с результатами другого, когда проводили пересадку печени молодого человека 20 лет старому (60 лет), что привело в течение 2-х лет к явным процессам глубокого омоложения, вплоть до восстановления седых волос в черные, улучшение зрения и многих иных внешне хорошо проявляемых признаков. Данный эксперимент показывает, что у человека стадийный сенилит это не необратимый процесс даже если первичная динамика его нисходит с верхних уровней регулировок и что определённая мощь поддержки реювенилизации идёт на верх с нижнего уровня, то есть геронтос* = единству взаимопроникающих сенесцента ⇆ сенилита. Реювенилизация онтостадийно зрелых и старых особей возможна и в мире животных.
Также правильность нашей теории подтверждают данные результатов по пересадке гипоталамусов молодых животных старым. Доказана возможность восстановления половых функций, гормонального цикла и веса матки и придатков у старых крыс при пересадке медиобазальных ядер гипоталамуса, что сопровождалось также рядом симптомов общего омоложения и активирования роста и общей жизнедеятельности.
Интерес представляют данные по непроизвольному самоомоложению.
Опишу пример японки Сэй Сенагон. Достигнув 75 лет, она почувствовала необъяснимые изменения в своём организме. Сначала у неё исчезла седина, и волосы приобрели былой блеск и черный цвет и неизвестно по какой причине вдруг стали резаться зубы! Затем стала разглаживаться кожа на теле и лице, мышцы приобрели былую эластичность, канули в небытие приступы остеохондроза и прочих старческих болячек, а через несколько лет её перестали узнавать подруги на улице, поскольку она помолодела лет на двадцать. Еще через некоторое время у неё возобновился менструальный цикл, новый муж утверждает, что Сэй выглядит не старше тридцати.
Удивительно и то, что через несколько лет она стала выглядеть гораздо моложе тридцатилетней женщины и серьезно опасается, что если процесс её омоложения сохранит нынешние темпы, то лет через 15 она превратится в 10 - 12-летнюю девочку!
Врачи полагают, что причиной чудесного омоложения женщины стали гормональные стимуляторы, которые вводились ей после перенесенной операции на печени. К сожалению, переподтверждений этому в последующем не было.
Если случай реален, то важно правильно дать оценку тому, что произошло в данном случае и является ли это самоомоложение результатом снятия процесса сенилита, то есть с гипоталамического уровня или это следствие клеточного снятия сенесцента, как это произошло в случае реювенилизации при пересадке молодой печени.
Аналогия принципа работы гипоталамуса с апикальным доминированием у растений
Предварительно напомню, что кроме апикального доминирования*, Гарбузов Г.А. ввёл понятие радиксального доминирования*, аналогом которого, очевидно, является эпифизарное доминирование*.
Чтобы понять и описать принцип работы гипоталамуса как регулирующей и движущей силы онтогенеза его можно сравнить с принципом регулировки созревания в растительном мире на примере апикальной меристемы дерева, в частности с той, которая связана с главной матрикальной = вершинной апикальной точкой роста, формирующей ствол. Это апикальная почка. В отличие от других апексов только она определяет полную соподчиненность всех остальных апексов на боковых ветвях, а также определяет начало наступления генеративной зрелости. Только вершинный апекс имеет достаточную для этого гормональную мощь, чтобы переподчинить себе все остальные точки роста. Гормональная мощь матрикального апекса складывается от суммы биомассы боковых ветвей, позволяющих в конечном итоге достигнуть требуемой мощи на главной почке. Дело в том, что апикальная почка настроена программативно так, что мощь её активности и роста многократно раз больше чем у всех остальных побегов и меристем. Конечно, это не обязательный принцип регулировок развития онтогенеза для всех растений, существует множество и других принципов, но именно по этому же принципу превалирования работает и гипоталамус. Энергия роста и гормональной мощи периферии обуславливает многократное превышение гормонов в главном апексе, который создаёт возможность для многократно усиленного стимула корней. Подчеркну, что на начальных этапах не сам апекс «тянет» периферию на созревание периферии, а наоборот. Включение программ на зрелость периферии возможно только при достижении определенного уровня гормонального профиля на главном апексе. Но для того, чтобы это произошло необходима достаточная масса и метаболическая мощь на периферии. Так происходит регулировка количественных изменений в качественные. За качественные изменения отвечает апекс. Сигнал на качественные перестройки периферия должна получить сверху, извне. В этом случае в апексе переключается ряд генетических программ на синтез новых регуляторов или многократного усиления тех, что уже были - гиперактивацию.
Угнетение вегетативного роста не позволяет созреть полноценно главному апексу.
У некоторых видов растений главенствует принцип не апикального доминирования, а - соматического. Некоторые виды из таких растений могут существовать на безлимитных принципах. То есть изнутри у них нет ничего, что мешало бы неограниченной их жизни. Это важно осознавать, с целью правильного поиска пути направления на воздействие корректировки онтогенеза.
Следует выделить, что существуют принципы апикального доминирования в виде условного и абсолютного апикального доминирования. При условном апикальном доминировании растение может многократно переоткладывать генеративные органы, а при абсолютном апикальном доминировании оно настолько сильно, что полностью заглушает возможность последующей перезакладки генеративных зачатков. Примером растения живущего по принципам условного доминирования является алое древовидное, смотрите ниже его фото, а примером абсолютного апикального доминирования является монокарпическая агава. Сюда же можно отнести и одно- или двухлетние растения, у которых тоже доминирует апикальная почка, но, по сути, это гипергенеративный тип доминирования, когда генеративная сторона развития полностью угнетает, исключает вегетативные возможности, то есть когда происходит полное подавление возобновления, инициации закладки всех типов последующих почек за счёт особого гормона абсцизовой кислоты. По сути, это явление полного запрограммированного самоуничтожения растения, это механизм полного феноптоза* всего организма. Если удалять эту цветочную почку, то растение может перейти в многолетники. Примером такой отмены двухгодичности может быть растение сельдерей, у которого можно удалять цветоносы и тем самым растение перевести в многолетник.
Гормон старения у растений имеет ли аналог в животном мире?
Итак, 1- и 2-х летний тип онтогенеза полностью подчинён главенствующей генеративной почке, мощь которой настолько сильна, что полностью блокирует инициирование любых последующих, как вегетативных почек, так и угнетение корневых зон роста, ответственных за выработку реювенилизирующего гормона цитокинин. Растение уходит в феноптоз* за счёт фитогормона стресса и старения абсцизовой кислоты. Вся регулировка онтогенеза растения исходит из одной точки. Если присмотреться к растению сельдерей, то можно заметить, что вместе с созреванием семян в его зонтике происходит одновременно пожелтение его листьев. А это элемент старения организма, причём наведённого сверху.

Рис. 7. На рис. а) видно что переход растения сельдерей на второй год в стадию образования семян вызывает у него пожелтение и быстрое старение листьев, а затем отмирание; на рис. б) внешний вид растения на первый год жизни, когда не закладываются семена, оно имеет зелёный молодой вид.
Такое старение, пожелтение листьев в разгаре лета аналогично поседению волос у человека. Это можно сказать навязанный, детерминируемый сверху тип старения, или точнее гипергенеративное апикальное доминирование. У растений нет явления всеобщего клеточного старения – сенесцента, то есть первичной направленности динамики старения с нижнего клеточного уровня. Следовательно, абсцизовая кислота, по сути, является гормоном старости и имеет полностью противоположное действие фитогормону молодости – цитокинину. Конечно, абсцизовая кислота отвечает не только за общее старение, но также и за локальные явления как апоптоз осенний у листьев и созревание плодов. Очевидно, локальный эффект старения у растений снимается более общим радиксальным эффектом омоложения. Следовательно, у растений имеется как гормон старения, так и гормон омоложения.
Если у растений гормоном стресса и старения является абсцизовая кислота, то у человека гормонами стресса является комплекс веществ, среди которых имеются как гормоны, так и нейромедиаторы. В частности, это сугубо гормон стресса кортизол и медиатор адреналин. К этой же группе следует отнести группу клеточных факторов провоспаления, проявляющих клеточный стресс. Каждый из них вносит свою лепту в общий плюрикомпонентный процесс. Причем, у человека такая регулировка старения, очевидно, двухэтажна, и даже трёхэтажна, как сверху со стороны эпифиза, гипоталамуса и комплекса нейротрофических факторов, так и снизу – со стороны клеточного уровня, то есть становления сенесцента. Похоже тут весомую значимость приобретает уровень сопровождения общего высшего уровня старения еще и нижним первичным клеточным старением. Седение (меланопения) является следствием двухуровневого процесса становления старения. Часть факторов имеет экзогенную природу (психологический или физиологический стресс, которые могут вести к преждевременному старению, то есть сугубо патогенез), а часть факторов имеет эндогенную природу и сопряжена с фазой онтогенеза, регулируемой через эпифиз-гипоталамус, ведущей в свою очередь к онтофизису, эпигенезу и как следствие сенесцент. Онтофизис ведёт к клеточному стрессу и затаскивает их в коммитированное состояние сенесцента. Коммитация происходит на уровне эпигенеза. Следовательно сенесцент без онтофизиса невозможен. Сенесцент сопряжен с Адаптационным Универсальным Клеточным Ответом (АУКО) — факторов клеточного стресса бывает множество, но механизмы их стресса и итоговые последствия тоже универсальны, сходны.
Для аналогии напомню про гормональную алопецию = облысение у мужчин, связанную с тестостероновой, то есть гормональной избыточностью. Эта проблема начинается у большинства мужчин после 40 лет. Облысение следует рассматривать как одно из множеств проявлений сенилита, когда организм выходит за рамки гормезиса по параметрам одного из гормонов. Этот выход свидетельствует, что организм вышел за пределы параметров оптимума, то есть туда, где начинается «открытый космос» и организм сам себя заводит туда, аналогично тому как это происходит у 1- или 2-х летних растений, где гиперапикальность заводит их к феноптозу или мортусу. У животных к этому же мортальному исходу ведёт гиперадальтус – аномально избыточная перезрелость. Этот гиперадальтус в определённой степени аналогичен репродуктивной перезрелости у растений - гипергенеративного апикального доминирования. У людей, очевидно, это можно обозначить как следствие гипергенеративного гипоталамического доминирования, которое является причиной механизмов реализации мортуса или феноптоза через анизомалии в виде атеросклероза, опухоли простаты как аденома, онкологии, саркопении… В растительном мире имеются виды с короткой длительностью жизни (1~2 года), средней (3~15 лет), долгоживущие (15~1000 лет) и с неограниченной. К последним относится корнеотпрысковая роща Тополя осинолистного, у которого нет механизмов выхода за пределы гормезиса и гиперапикального доминирования. Человеческий организм следует отнеси в группу средней продолжительности жизни, так как их потенциал заранее ограничен гипоталамическими сроками выхода организма за пределы гормезиса. Все признаки анизомалий у него начинают проявляться в 35 лет и доходят до критических в 65~100 лет.
Очевидно, гиперадальтус является причиной неизбежного образования с возрастом аденомы простаты. Облысение – результат сдвига сугубо гормонального фона и является следствием гипергипоталамического доминирования, а не клеточного сенесцента*. Очевидно, у животных тоже имеется такой насильно навязанный фактор старения и таким «гормоном старения» является новый профиль или изменение возрастного соотношения гормонов и трофических нейромедиаторов, которые триггерно сцеплены с программой гомойотопии*, и которая их провоцирует на усиление. Конкретного гормона старения или молодости у животных не известно, но феномен старения (сенилит*) или смена одного фенотипа молодости на фенотипы зрелости или старости может стать проявлением в изменении гормонального профиля, пейзажа по целой группе гормонов, в отличие от растений, где регулирование происходит преимущественно за счет преобладания гормона молодости или старости. Например, избыток тестостерона и кортизола на фоне недостатка мелатонина? Но, тем не менее, при кастрации человека снижение уровня тестостерона не гарантирует существенное отодвижение сенилита организма. Уровень мелатонина при этом не обязательно поднимется. Значит генеративной доминанты над ним нет? Онтогенез человека плюрифакторен? В эту систему вмешивается фактор сенесцента?
Где в организме указано кому сколько жить?
Отмечу, что у растений с 1- или 2-х летним типом онтогенеза он регулируется с помощью генеративных органов, что в свою очередь обуславливает программу стресса и истощения всего организма через посредство гормона стресса абсцизовой кислоты.
Онтогенез 1- и 2-х летних растений определяет принцип генеративного доминирования. Так у 1-летней пшеницы или 2-х летнего сельдерея жизненный цикл определяет гормон старения – абсцизовая кислота (АК), которая образуется в созревающих семенах, причем так мощно, что подавляет все жизненные показатели растения и выводит его полностью на феноптоз. У агавы жизненный цикл 15 лет и у данного монокарпического растения мощный цветонос становится фактором самоуничтожения растения тоже за счёт АК. Этот жизненный цикл у агавы можно продлить до 100 лет, если удалять цветонос. У многих бамбуков жизненный цикл около 100 – 200 лет, затем наступает резко фаза зрелости и вся роща начинает колоситься, зацветать, образуя множество семян, при этом сама в тот же год полностью гибнет. Очевидно, все эти 100-200 лет бамбук рос вегетативно и всё это время цитокининовый профиль был преобладающим, что продлевало на многие годы и столетия онтогенез. В данном случае можно говорить о вегетативном профиле, или о принципе вегетативного доминирования, когда все эти годы происходит превалирование цитокинина – гормона молодости над всеми другими гормонами. У сосны остистой с жизненным циклом не менее 4800 лет тоже её жизнестойкость обеспечивает особенность её корней, которые с избытком вырабатывают цитокинин и тем самым дают возможность оживления самым заглушенным побегам и почкам. Причём у деревьев мафусаилов-долгожителей может тоже проявляться ежегодно генеративное доминирование с образованием огромной массы стробил, то есть женских и мужских шишек, что ведёт к увеличению уровня АК - гормона старости, с последующим усыханием ветвей. Но, тем не менее, дерево умеет преодолевать эту локальную старость ветвей за счёт цитокининового омоложения из корней. То есть гормон старости АК воздействует мозаично не на все части растения, а только на наиболее приближенные; точно также и цитокинин достаёт и воздействует только на часть побегов, но при этом часть побегов ему остаются недоступными, это чаще всего побеги отплодоносившие, также плагиотропные, то есть нижние, свисающие вниз с ограниченным потреблением нутриентов и гормонов. Они то в первую очередь и усыхают. Такая мозаичная регулировка онтогенеза присуща большей части древесных растений. Это так называемый промежуточный тип между вегетативным и генеративным доминированием. Онтогенез таких растений может продолжаться десятилетиями и столетиями, когда происходит уравновешенное балансирование между вегетативным и генеративным доминированием, то есть процессами старения и омоложения. Но, в любом случае, продолжительность онтогенеза как дерева в целом, так и отдельных его ветвей определяют гормоны старения и молодости, а точнее их соотношения.
В ряде случаев можно говорить о промежуточном доминировании над определёнными побегами гормона молодости – цитокинина, тогда как у части побегов происходит доминирование гормонов старости и их усыхания за счет доминирования сугубо в них абсцизовой кислоты. Часть побегов дерева являются стадийно старыми, а часть – стадийно молодыми. Постепенно этот фон меняется в сторону всё большего преобладания АК и уменьшения значимости гормона ЦК и все больше и больше ветвей уходит на усыхание. Так дерево стареет целиком. Гормоны старения постепенно становятся преобладающими над гормонами омоложения.
Особенности старения на региональном уровне у животных.
Очевидно, аналогичная картина имеется и в организме человека, что приводит к постепенному угнетению ряда стволовых клеток, дряхлению и склеротизации ткани. Только в отличие от растений, где стареют определенные ветви или зоны роста, в организме человека таковыми являются точки самовосстановления, имеющиеся повсеместно во всех тканях, в которых происходит балансирование между старением с его апоптозом части клеток и одновременного обновления новых линий клеток из стволовых. Это так называемая полная реювенилизация, которая наблюдается у простейших видов животных, например гидр, планарий и других. Но у высших животных всё усложняется вмешиванием в этот процесс полного самообновления еще и сенесцентом клеток.
У рыбы горбуши, также как и у растений с генеративным принципом доминирования регулировки онтогенеза, аналогично таким же мортальным фактором являются гонады, посредством многократного и фатального гиперфункционирования гипоталамо-гипофизарной системы, роль которых у неё становится доминирующей.
Таким образом, весь мир животных и растений в филогенезе разработал множество правил для проявления различных типов онтогенеза, где управление им может идти как с верхних уровней, так и с нижних. Например, управление сверху происходит за счёт доминирования верхних уровней регулировок в гипоталамусе или на главных апексах растений. Но эти же параметры онтогенеза могут регулироваться не обязательно на верхних этажах регулировок, возможны ситуации когда онтогенез определяет Жизненная Сила низов. Так, например, у дерева сосна остистая, которая живёт тысячелетия, тонус жизненной силы определяют не апикальные точки роста, а гормональная сила корней, их сила превалирует над силой верхушечных апексов. В этом случае даже «погибающие» части дерева самовозрождаются «как из пепла» в виде отрастания новых побегов из глубоко заглушенных в норме брахиальных почек (в пазухе хвоинки или листочка). Параметры онтогенеза очень лабильны и могут базироваться на совершенно противоположно направленных принципах, что позволяет природе маневрировать от 1-летнего типа онтогенеза до тысячелетий.
Существуют виды растений, у которых угнетение вегетативного роста является фактором включения программ на ускоренное созревание. Это так называемые «скороспелки», у которых укорачивается онтогенез за счёт укорочения ювенильной фазы и ускорения фазы сенилита. Причём такая скороспелость может быть как эндогенно предопределённой (например у однолетних растений), так и связана с внешними условиями. Например, дерево лиственница, которая в норме начинает активно плодоносить в природе на 20-м году жизни, но при посадке в каменистую бедную почву может заплодоносить на 5 году жизни.
Аналогично у человека также можно укоротить ювенильную фазу детства и юности и ускорить наступление последующих фаз.
Функция матрикального апекса* многогранна. Это означает, что апикальная точка определяет как морфогенез других частей дерева и его ветвей, так и сроки перехода от сугубо вегетативной фазы и готовности к наступлению генеративной зрелости. Например, у хвойных растений это сроки закладки первых стробил = шишек.
Онтодианность длительности онтогенеза.
Таким образом, можно отметить, что как в растительном, так и животном мире имеются виды, где одни из них проявляют короткожительство, а другие – долгожительство, которое обеспечено у каждого конкретного вида своим механизмом регулировки онтогенеза. В связи с этим нами предложено ввести понятие онтодианность*, которое означает, что у каждого вида длительность жизни изначально коммитирована, то есть генетически запрограммирована. Термин созадан по аналогии с циркадианность, когда ритмы жизни связаны не с внешними факторами, а внутренними. Поэтому можно говорить о растениях с 2-х летней онтодианностью, 3-х летней, многолетней.
Механизм перехода апекса из вегетативного в генеративное состояние.
Срок готовности зрелости определяется по мощи происходящих гормональных процессов на периферии, то есть готовность назначает не сам по себе матрикальный апекс, а через его связь с периферией; мощь метаболических процессов и гормонального фона на периферии запускают (экспрессируют готовые к этому гены) в апексе заблокированные до этого механизмы переключения программ на фазу генеративной зрелости. Низы определяют сроки перестройки в верхах, а реализация гормональной зрелости верхов определяет нисходящие сигналы, команды на зрелость низов, то есть разблокирование программ морфогенетических перестроек обеспечивающих репродуктивную зрелость. Вся система развивается динамически со всех сторон, одно усиливает развитие другого. Сигнал к запуску генеративных программ определяет размер апекса, прежде всего количество меристематических клеток и тканей, пока не достигнет критической массы, дальнейшее развитие репродуктивного этапа не подключается. Следовательно, включение пускового механизма зависит от степени чувствительности генотипа к сигналу и онтогенетического состояния растения. Также известна азотно-углеродная (трофическая) теория регуляции цветения. Если в растении много соединений азота и мало соединений углерода, оно будет расти, а если углеводы преобладают над азотом, то растение цветет. Очевидно, это является ключом к пониманию регулировки переключения программ: азотистые соединения>углеводы.
По концентрации азота растение «оценивает работу» корневой системы, а значит и уровень цитокининов (гормона молодости и вегетации), а по сахарам — «работу» листового аппарата. Если листовой аппарат «работает хорошо», то много ауксинов, высокие концентрации которого указывают на возможность плодоношения. Но это предрасполагает, даёт возможность, но начало плодоношения определяет степень чувствительности генотипа, который может отодвинуть это начало на год или на десятки лет. Также если работа корня слишком интенсивна, то нужно восстановить баланс между активностью корневой системы и площадью листового аппарата. Растение образует много листьев (вегетирует), а когда баланс восстановится, образует цветки и плоды.
Аналогично похожие механизмы регулировки перехода из состояния юности в генеративное через гипоталамус у организмов животных: адениновые вещества, аминокислоты>углеводы и жиры. Но стимулировать рост в организме, анаболические процессы неограниченно тоже нельзя. Это означает, что азотистые соединения должны быть на нижнем минимуме самодостаточности для самоподдержания, чтобы не провоцировать преобладание анаболизма над катаболизма, но держать их в балансе и всё это на фоне возможно допустимого минимума углеводов, что должно теоретически сдерживать наступление зрелости как соматических тканей, так и гипоталамуса.
Низы настроены находиться в состоянии непрерывного роста, чтобы в заданное время приблизиться к критическому порогу набора нужной биомассы, идёт самонаращивание биомассы, что обеспечивает возможность набрать силу и нужный рост одновременно и главному апексу. При этом главный апекс настроен так, что скорость гормонального роста в нём не пропорционально многократно раз выше чем в других апексах. Это условие его доминантности. Этот рост продолжается до тех пор пока не достигнет критических параметров по гормонам и метаболому. Верхняя апексная точка является детектором определяющим достижение требуемых параметров, что позволяет экспрессировать новую зону генов и обеспечиваемых ими программ на генеративное созревание и производство гормонов нужных для этого.
Из этого следует вывод, что не верх определяет статус низов, а наоборот. Верх только координирует сроки перехода на новые этапы. Апекс это чувствительная система, которая работает по принципу «наполнения» ёмкости, которая пока не наполнится до верху водой, не откроется поплавковый механизм переключения и закрытия напора воды из крана. У растения этому аналогично свободное вегетирование, которое происходит «самотёком» и при достижении определённых уровней вегетативная фаза притормаживается и включается генеративная.
Аналогичное происходит и в гипоталамусе. Но, тем не менее, ряд авторов [1] утверждает, что переключение в гипоталамусе происходит на других принципах и выдвигают концепцию, что эта функция переключения на фазу половой зрелости находится в самом гипоталамусе, в котором есть система «Тэнд» – отсчёта времени, а гипоталамус называют гомеостатом времени. Эти авторы предложили ввести понятие о гипоталамусе как регулятора эндогенного времени = Тэнд. Но такая трактовка означает, что это система, которая развивается и работает сама в себе, не корректируется общим гормональным фоном. Это означает, что функция регулятора времени изначально должна быть коммитирована генетически, то есть изначально включенной программой для данного типа клеток. Это очевидная ошибка. Ведь в этом случае эта функция корректировки сроков этапов онтогенеза гипоталамуса работает сугубо сама по себе и не работает совместно с динамикой происходящей на нижних этажах подчиняемых ему. Ниже мы рассмотрим как на созревание гипоталамуса и степень его чувствительности к сигналингу влияет влияет уровень мелатонина в эпифизе. Он и является драйвером онтогенеза. Напомню, что матрикальный апекс у дерева созревает, набирает мощь вместе с ростом всего дерева, а не сам по себе. Да этот апекс становится доминантной структурой, но при этом зависящий от низов. Вся система дерева развивается динамично во всех направлениях и одно не может развиваться без другого. Точно также должно происходить и в организме человека, где тоже происходит длительная фаза роста = вегетации, становления. Не следует путать способность системы регулировок в отношении перехода на новые фазы развития к внутренним перестройкам как автономный процесс. Гипоталамус находится в постоянной динамике, развитии и созревании. Это структура многоядерная, мультицентровая, но в итоге даёт общий единый результат по направленности и срокам развития организма. И не может какая-то одна из его функций работать сама по себе
Проанализируем как происходит регулировка фазами онтогенеза у деревьев, у которых организован механизм так называемого апикального доминирования, которому подчинён контроль за управлением всей гормональной системой дерева. Гипоталамус, по сути, тоже можно сравнить с матрикальным апексом, а значит он тоже как дирижёр руководит всей гормональной системой организма. Гипоталамус тоже матрикален, стоит во главе гормональных многих осей, с ним согласовывается и от него отталкивается, синхронизируется всё развитие. Начало генеративного развития, зрелости дерева начинается когда матрикальный апекс достигнет своего критического предела мощи, достаточной чтобы запустить соответствующие на это рецепторные гены, которые настроены на экспрессию при достаточной мощи растущей ткани и объёма гормонов, чтобы «распаковать» нужные гены, сделать их чувствительными к пробуждению и переходу на новую программу развития, а значит становления нового этапа онтогенеза. Для того, чтобы дерево заплодоносило, апекс должен приобрести готовность к этому, созреть. Например у агавы, которая даёт цветонос один раз в жизни, и относится к монокарпикам, это событие происходит обычно через 15 лет. Но если у этой агавы декапитировать (удалять) цветонос, то она может жить до 100 лет. В данном случае гиперзрелая апикальная точка угнетает все ткани с усыханием и гибелью всех листьев, а все соки и энергию отдаёт цветоносу. Апекс определяет длительность жизни и начало генеративной зрелости.
Но у агавы есть еще возможность к вегетативному саморазмножению, путём образования ювенильных боковых точек роста в основании, что ведёт к появлению множества ювенильных отводков, «деток». Такая настройка надолго может тормозить образование генеративного цветоноса. Вегетативная фаза и ювенильность преобладают над генеративной и сенилитной. Апикальное доминирование апекса здесь ослаблено в сторону усиления возможностей вегетативного соматического доминирования и тем самым поддержания статуса ювенильности. Гормональный фон таких агав имеет крен в сторону настроенности на ювенильное существование. Это означает, что программа на поддержание уровня стволовых клеток с более высокой потентностью превалирует над программой ведущей к генеративной зрелости.

Рис. 8. Агава с множеством боковых отводков, «деток»; такая агава не сможет дать цветонос и сохраняет своё ювенильное состояние, неотению. Детки у неё закладываются из спящих почек, имеющих максимально высокую степень потентности стволовых клеток (плюри- и тотипотентность). Такое состояние неограниченной неотении у агавы можно вызвать искусственно, удаляя цветонос, или в случае генетической трансформации = коммитации на непрерывное вегетирование. Например, такая коммитированность появляется у алое, когда новые побеги образуются из брахиальных почек (пазушных почек у листьев), но они имеют меньшую степень потентности стволовых клеток, более зрелы и способны на следующий год дать цветонос.
В организме животных тоже можно найти такую аналогию по превалированию программ ювенильности и поддержания высокого уровня стволовых клеток с более высокой потентностью, способных не только обеспечить репарацию ткани, то есть замены в ней старых клеток на более молодые, но и на регенерацию ткани в случае необходимости. Если бы в организме человека более активно работали стволовые клетки с более высокой потентностью, то тем самым они бы подавляли активность гипоталамуса, притормаживая его перезревание, ведущее к матуре, адальтусу и сенилиту. Вспомним эксперимент с мышами, которых ввели в парабиоз, когда молодая особь омолаживала старую. Следовательно, не сам по себе гипоталамус ведёт к матуре и сенилиту, а состояние всей системы в единстве. Также эксперимент с парабиозом подтверждает, что в гипоталамусе нет счётчика времени «Тенд».
Точно также у рыбы горбуши наступление зрелости и перезрелости гипоталамуса определяет её способность давать потомство один раз в жизни на девятом году. Монокарпизм горбуши аналогичен доминированию репродуктивного апекса у монокарпической агавы. Монокарпизм у этой рыбы обуславливает особенность работы гипоталамуса, который задаёт особый скоротечный тип адальтуса из-за гиперизбыточности гонадных гормонов, а также особого типа сенилита, с жестким механизмом мортальности. Если у горбуши удалить гонады, то она может прожить 28 лет и не стареть.
Но природа сама способна производить декапитацию, то есть когда эта программа не становится главенствующей и окончательной, а за ней могут повторяться следующие циклы роста и генеративной зрелости. Например, у алое древовидного, другого вида растений из того же семейства, что и агава, но после цветения и плодоношения оно не погибает, а образует новую боковую точку роста.

Рис. 9. Алое в виде дерева. У растения нет главного матрикального апекса, а также нет гиперапикального доминирования и монокарпизма. В принципе такое растение могло бы существовать неограниченно, если бы не мешали внешние факторы нужные для непрерывного роста. На вершинах некоторых побегов видны многочисленные цветоносы. Растение полиапексное и поликарпик.
У такой агавы образуется ствол, ветви, на которых образуются такие же мясистые розетки жирных листьев, на концах которых образуются цветоносные побеги. Цветоносы у них мелкие и не ведут к гиперапикальному доминированию, то есть не заглушают последующие точки роста. Поэтому алое после цветения не гибнет, здесь нет принципа генеративного доминирования, то есть принцип абсолютизма, когда организм ради превалирования генеративных возможностей жертвует всеми остальными точками роста, полностью заглушая их.
А теперь давайте посмотрим на этот же принцип долгожительства у деревьев на примере хвойного дерева сосна остистая, способной доживать до 5000 тысяч лет.

Рис. 10. Сосна остистая, возраст которой насчитывают 4850 лет, но Жизненная Сила не покидает её.
Дерево представляет собой гигантскую махину ствола, которое, по сути, представляет собой остов из сухой части ствола с отсутствием верхушечного матрикального побега, но, тем не менее, сохраняющего силу роста у части боковых ветвей, которые морфологически не проявляют угнетенный или сенилитный тип роста. По сути, это чередование мертвых частей ствола и множества толстых сухих ветвей. Явно, силу нового роста от боковых ветвей даёт дереву мощь его апексов на корнях, а точнее мощная живучая корневая система. Корни дерева заставляют его продолжать жизнь, именно они берут на себя главенство по проявлению особенностей онтогенеза этого дерева. В данном случае следует говорить о корневом апикальном доминировании, а не о матрикальном. Этот тип онтогенеза нами предложено обозначить как радиксальное (корневое) апикальное доминирование. Особенность этого типа онтогенеза – способность сохранять, поддерживать высокий уровень молодости, высочайшую способность к регенерации. Только хорошо развитые корни способны вырабатывать преобладающее количество цитокинина, обеспечивающего неограниченную регенерацию, а также сдерживать доминирующую избыточность генеративной зрелости.
Оказывается в мире растений такой тип неограниченного радиксального онтогенеза встречается довольно часто и, кроме того, который позволяет не только пролонгировать жизнь, но и длительно, вплоть до бесконечности удерживать признаки ювенильности. Никаких принципиальных препятствий этому нет. Таковым является например плющ. Особенностью этого растения-лианы является то, что на одной особи одновременно могут находиться и онтостадийно зрелые ветви с плодами и ювенильные побеги. Зрелые с плодами ветви располагаются в верхней части растения, где больше света и доминирует работа ауксина, а нижние с ювенильной формой листвы расположены внизу в более затемнённых местах, где имеется в пазухе каждой брахиальной почки зачатки корней, а это означает локальное радиксальное доминирование за счёт избытка цитокинина. Также это радиксальное доминирование обеспечивает неограниченное существование вегетативной роще тополя осиноподобного, бамбуковым рощам, живущим сотни лет, или деревьям мафусаилам как секвойядендрон и другим. Все растения, которые могут перезакладывать свои части в новом месте способны существовать неограниченно. Для этого им достаточно иметь способность закладывать спящие почки. Но не все деревья имеют такую способность. Например, у сосны остистой нет такой способности давать поросль, но зато она способна легко пробуждать адвентивные почки на ветвях. Это обеспечивает ей высочайшую живучесть. В свою очередь секвойядендрон (мамонтовое дерево) не имеет способности закладывать спящие почки в основании ствола, тогда как эта способность имеется у секвойи вечнозеленой, что позволяет ей легко отбивать новую крону дерева при пожарах и тем самым продолжить своё существование. Секвойядендрон при пожарах погибнет, но если их не будет, то тоже благодаря своей особой мощной корневой системе способен жить несколько тысяч лет.
Фаза ювенильности может быть растянута у растения вплоть до бесконечности, но может быть прервана в любой момент пока не получит достаточный уровень ауксинов, которые вырабатываются в условиях фотоиндукции, то есть на свету.

Рис. 11. Ювенильные и взрослые плодоносящие побеги взятые с одного растения.
Ювенильные побеги у плюща никогда не дают образование плодов. Плоды образуются только на морфологически зрелых укороченных ветвях. У ювенильных побегов листья вырезанные как у клёна и имеется большое количество корневых зачатков (ювенильной меристемы, мезенхимальной ткани), побеги вытянуты, легко укореняются, а у зрелых – листья овальные, на верхушечных апексах закладываются плодовые почки, которые тормозят рост в длину побега. Побеги укорочены, что говорит об угнетении гормона роста гиббереллина. Гиббереллин можно признать аналогом гормона роста соматропина у животных. Гиббереллин и соматропин всегда сопровождают ювенильную фазу вегетирования. Но в тоже время гиббереллин нужен для вегетирования, но не обязателен для ювенальности, например рост из брахиальных почек ослаблен, у спящих почек и порослевых побегов он может быть сильный, но ювенильность вполне может происходить без сопровождающего роста. Это означает, что в организме человека нужно найти гормон ювенильности, который бы работал сам по себе без гормона роста. Поэтому принцип гомойотопности не является помехой для сохранения ювенильности, гормоны регулирующие эти процессы и нужные программы разные. У плюща на верхушечных побегах наблюдается выраженная продвинутая онтостадийность.
Важно обратить внимание на особенности развития плюща, в частности на то, что верхушечные побеги прекращают рост и проявляют апикальное доминирование и высшую степень стадийной зрелости, тогда как нижние побеги имеют все признаки ювенильности. На одном растении одновременно находятся и апикальное доминирование и радиксальное доминирование. Прекращение роста можно сравнить с механизмом гомойотопии у организмов животных, которая наступает после половой зрелости. Торможение роста у животных = гомойотопия – аналог апикального доминирования и высшей степени стадийной зрелости. Аналогичный элемент радиксального доминирования (ювенильности) в организме животного на постпубертатной стадии полностью заблокирован. Если в организме растений гормон, который отвечает за ювенильность цитокинин известен, то в организме животных – он еще не определён до конца, но ясно одно, что он тоже обязательно есть.
Если в нашу задачу входит реювенилизация организма, как это легко происходит у плюща, то надо задействовать гормон противодействующий стадийной зрелости. Следует обратить внимание, что у плюща этот гормон юности открывает одновременно и функцию роста растения через ауксины и гибберелины. Можно было бы предположить, что у гомойотопных видов животных эта реювенилизация откроет заодно и фазу роста организма, что противоречит его жизненным принципам. Но не следует забывать, что рост инициируют гормоны ауксины, а реювенилизацию – цитокинины. Следовательно, усиление в организме животного гормонов реювенилизаторов = аналогов цитокина, не усилит вероятность очередной вспышки роста ткани, а всего лишь обеспечит самовосстановление стволовых клеток.
Тем не менее, эволюция животного мира пошла по пути создания таких организмов, у которых исключена возможность повторной перезакладки новой мощной мезенхимной ткани, а взамен оставлены только нижние порядки стволовых клеток как прогениторы и им подобные, позволяющие производить замену клеток, но онтостадийно зрелых. Это означает, что в организмах животных предусмотрена только разовая закладка эмбриональной ткани с тотипотентными свойствами, тогда как у растений эта возможность не отменяется в онтогенезе и они легко могут заново переотложить отпочковавшиеся растения или воссоздать, регенерировать утраченные апексы, ветки и корни… Кроме того, у животных полностью исключена возможность реювенилизации тканей с помощью некого общего универсального для всех тканей и видов растений гормона молодости.
В чём сходство и различие точек самовосстановления у животных с зонами роста у растений?
У растений зоны роста во всех почках являются одновременно и зонами самообновления, которым подчинён рост камбия сосудов, флоэмы. Растение самоомолаживается ростом во вне. Они же имеют максимальную степень свободы, потентности по регенерации любого своего органа в любой ткани, например на корнях могут заложиться почки, из которых отрастут новые побеги и ствол, а на листьях, побегах и стволах могут заложиться корни и т.п. В животном мире тоже есть точки самовозобновления, но практически с минимальными потенциями на регенерацию, кроме того работают они на других принципах и программах. Главная особенность в том, что такая точка самообновления находится внутри и не может разрастаться, а обязательно должна соблюдать постоянство размеров, что означает необходимость замены старых клеточных линий на новые, когда сколько клеток было подвержено элиминации – столько же должно прийти на их место! Здесь нет единого универсального для всех тканей ювенилизирующего гормона, а есть ряд специфичных гормонов, присущих для конкретных тканей. Эти точки являются аналогами зон роста у растений. Та обновляемость клеток, которая сохранилась, вполне может регулироваться не за счёт гормонов глубокой реювенилизации и дедифференциации, а за счёт гормонов роста. Возможно, аналог гормону цитокинин у животных имеется только в эмбриональной стадии бластулы.
Также отмечу, что у концепции гомойотопии = лимитированного роста, как ограничивающего фактора на реювенилизацию, имеются факты идущие ей вразрез. Например, жизнь голых землекопов основана на принципах неотении, продленной молодости, но при этом их размеры и рост не отличаются от мышей, продолжительность жизни которых в 10 раз меньше. Решающим здесь являются особенности работы их гипоталамуса. Похоже что с учётом того, что они ночные животные, проживающие под землей, эпифиз у них с возрастом не прекращает свою работу, а значит выработка мелатонина не останавливается, что не позволяет перейти гипоталамусу на принцип доминирования и гиперзрелости.
Выше мы обсуждали пример с плющом, который показывает, что в растительном мире реально имеется возможность разблокировки и осуществления как полной реювенилизации всего организма, так и его бесконечной регенерации. Для этого им нужно иметь возвратные механизмы переключения на прежние исходные программы и создавать особый гормональный фон. Это означает, что онтогенез у растений может двигаться как вперёд, так и назад и имеет принципиальную возможность продлеваться неограниченно. У животных гомойотопных* такой базы нет и поэтому её надо искусственно создавать. Эмбриогенез у них однократный.
Так у ряда растений особенность роста их корней даёт дополнительную гормональную силу, которая позволяет ей пробиваться через усыхающие ветви с затухающим ростом на всех апексах и закладывать всё новые и новые боковые ветви. Если посмотреть на общее состояние дерева-долгожителя, то бросается в глаза огромная масса полностью отмерших частей дерева. Дело в том, что у растений нет механизма и принципа очистки изнутри от погибших клеток и частей. Да, у растений есть механизм апоптоза, который, например регулирует опадение листьев или веткопад, но это возможно только на внешней части организма и никак не контролируется изнутри.
Дерево может существовать только за счёт мощного израстания наружу новых почек и побегов. То есть они образуются только за пределами предшествующей ткани. Новое здесь не происходит за счет полного ликвидирования старого. Это придаёт особый тип старения растению, когда сочетается дряхлость, развалины умерших частей остова ствола и ветвей и мощь нарастания новых ветвей. Эту мощь самообновления, то есть Жизненную Силу придаёт дереву мощь корневой системы. Её сила жизни и гормональный фон (гормоны молодости растения цитокинины) заставляют пробиваться новым брахиальным побегам в кроне и постоянно их обновлять. Корни дают силу молодости стареющим с затухающим ростом ветвям, заставляют пробуждаться и проявлять новую энергию роста. Но происходит это уже преимущественно не за счёт восстановления роста в дряхлеющих апикальных почках или матрикальной главной, а за счёт отрастания брахиальных (пазушных) почек, которые более чувствительны к цитокинину чем к ауксинам. Значение цитокининов начинает преобладать над значением ауксинов. Апикальные почки в норме сами себя обеспечивают энергией роста за счет аттракции (натягивания) на себя гормонов и нутриентов, тогда как пазушные почки или спящие ювенильные почки получают больший стимул энергии роста из корней, но при условии угнетения апексов. У старых деревьев апикальное доминирование кроновых почек резко ослабевает, а снятие этого доминирования позволяет разблокироваться боковым почкам. Это их принципиальная биологическая особенность. Растение переходит на принцип существования за счёт превалирования мощи стволовых клеток или почек второго порядка, в том числе и корневых. В норме они являются соподчинёнными точками-мишенями для регулировки сверху, но когда апексная регулировка отменяется, исчерпывает себя, на себя берут эту функцию продолжения роста другие типы точек роста. Соподчиненность точек роста второго порядка снимается, они растормаживаются и становятся основными.
В принципе мощь Жизненной Силы корней можно было бы приравнять к мезенхимным стволовым клеткам и зависящих от неё прогениторным концевым стволовым клеткам в организме животных. Но к сожалению, в отличие сверх развитой мезенхимы корней данной сосны, у человека эта мезенхима не получила должного развития и регулировка идёт преимущественно только за счет прогениторов с низкой степенью потентности. Если бы животные имели бы такую же мощь на постоянное самообновление ткани, как это происходит на примере сосны остистой, то это бы позволило бы организму животного получать такую же непрерывную мощь на самообновление, то есть сохранения равновесия между процессами внутренней самоликвидации за счет апоптоза и регенеративными процессами. Таким образом могут создаться условия саморегулировки поддержания непрерывности циклов репарации и регенерации. Этот же процесс содействовал бы запуску механизмов по возврату клеток от сенесцента к ювентальности. Если такого равновесия нет, идёт односторонний процесс, когда ткань стареет. Конечно, для этого у организма должна быть хорошо проработанная система самоочистки от отмирающих частей, в которой главенствующую роль должны сыграть механизмы апоптоза, аутофагии и аутолиза.
А теперь давайте сравним облик тысячелетнего дряхлого дерева с внешним типажом старого человека.

Рис. 12. В крайней степени проявленный фенотип старческого облика у человека связан с качественными повсеместными признаками дряхлости на уровне всех клеток и тканей.
На рис. 12 представлена особенность фенотипии (внешнего облика) старого человека, проявленная в крайней степени. Эти признаки дряхлости проявляются в виде: меланопении (поседение волос), алопеции (облысении), саркопении (мышечная слабость), дерматопении (кожная вялость), гиперкератоз старческий (грубое утолщение кожи), гиперлипофусциноз (гиперпигментация, кожные пятна от избыточного отложения липофусцина, который занимает 25-50% объема клеточной цитоплазмы, что и обуславливает бурый или землистый оттенок кожи), гиперругоз кожи (морщины), гиперадальтус (гормональная тестостероновая перезрелость, гирсутизм бороды, избыточный рост волос бровей, но алопеция на голове), акромегалия – избыточный рост подбородка и мочки ушей. Рассматривая весь приведённый здесь симптомокомплекс фенотипа старости следует определиться является ли он следствием гормональных верхних перестроек, или следствием – сенесцента? Является очевидным, то что часть из них является прямым следствием гормональных перестроек, например алопеция. Но другие симптомы являются косвенными, вторичными, опосредованными, реализуясь в общей динамике становления организма через сенесцент. После реализации сенсцента, как результата онтофизиса и эпигенеза, становится возможным то, что сенесцент тоже становится причиной и механизмом для проявления сенесцент-зависимой симптоматики сенилита. Последствия сенесцента – симптоматичны, а сенилита – этиологичны. Конечно, более правильный подход через устранение этиологии. Например, гиперлипофусциноз, то есть отложение липофусцина в постмитотических (неделящихся, застрявших на фазе G0 - покоя) клетках - нейронах, кардиомиоцитах, волокнах, эпителиальных клетках, пигмента сетчатки. Является ли это следствием или причиной сенесцента? В статье «Гипоталамус – верховный драйвер онтогенеза» [10] автор приводит обоснование того, что сенесцент это следствие онтостадийного продвижения к сенилиту. Таковым является то, что у долгоживущих типов животных и человека процессы сенесцента, анизомалийные перестройки и анизомалические патологии проявляются намного в более старшем возрасте – после 40-60 лет, тогда как у короткоживущих животных (мыши) тот же самый спектр анизомалийных болезней, но сенилит проявляются после 2 лет. Чем глубже продвижение по этапам онтогенеза, тем жестче сенесцент, так как организм всё дальше и дальше выходит за горизонты гормезиса и гомеостазов, в условиях усиления до крайности гормонального гипердоминирования. Сенесцент коммитирован, то есть фиксирован.
Итак, все качественные признаки старения у человека резко отличаются от признаков старения (дряхлости) у старого дерева сосна остистая, у которой на локальном уровне побегов нет признаков старости, что подтверждает что у растений отсутствует клеточный сенесцент, а есть только сенилит.
Сравнение этого облика старого человека с лицом юноши показывает поразительные возрастные изменения качественного и количественного характера. Они указывают на то, что главный орган ответственный за поддержание гормезиса, то есть гипоталамус, у старика давно вышел за пределы оптимума, а клетки работают за пределами гомеостазов.

Рис. 13. Фенотип юношества имеет колоссальные отличия от фенотипа сенилита.
У человека роль апексов отчасти играют прогениторные и др. стволовые клетки. Они затухают аналогично тому, как затухают апексы у старых деревьев. В связи с затуханием роли гипоталамуса, то есть повышения его резистентности = нечувствительности (аналог верховной матрикальности) происходит затухание, угнетение прогениторов – стволовых клеток второго порядка (аналог соподчиненных апексов). Но у дерева сосна остистая открыт канал для подпитывания мощной Жизненной Силы посредством сохранения мезенхимных клеток на корнях. Они дают «соки», которые оживляют и позволяют пробиваться через старые, полу усыхающие ткани и дряхлеющие почки новым, но уже не за счёт продолжения роста главных апексов, а за счёт прорастания брахиальных и спящих почек. Такие почки имеют меньшую степень генеративной зрелости и побеги от них не дают так быстро обилие шишек, что меньше истощает растение в целом.
Цитокинины – гормон молодости и регенерации у растений вырабатываются в корнях.
Если сосне остистой возвращают жизненную силу особенности её мощной корневой системы, то почему этого явления не наблюдается у других видов сосен и они попросту при болезнях и с возрастом усыхают и не борются за жизнь? Очевидно, это связано с наличием у сосны остистой особо развитой мощи корневой системы с обилием зачатков мезенхимы, то есть более ювенильной, она меньше поддаётся сигналу на гормональное созревание идущего из главных апексов, то есть это сигнал на перезревание и торможение, угнетение. Но в тоже время корневая мезенхима способна создавать обилие ювенильных гормонов способных оживлять даже усыхающие ветви, в частности цитокинины. Больше всего цитокинины вырабатываются на апексах корней. Если ауксины синтезируются в апексе побега, то цитокинины на кончиках апексов корня. Ауксин транспортируется по растению сверху вниз и активно, а цитокинин - снизу вверх и пассивно. Чем сильнее корни, тем больше гормона молодости и жизни, а при доминировании в росте корневой системы они смогут оказать существенное влияние на увеличение продолжительности жизни ствола и кроны дерева. Смотрите пример их омолаживающего действия на фото ниже, где стареющие и желтеющие листья обрабатывали этим гормоном и происходила их реювенилизация.
Очевидно, в организме животных тоже должен быть аналогичный клеточный гормон ювенильности, который может преодолеть клеточный сенесцент и митохондриальную дисфункцию.
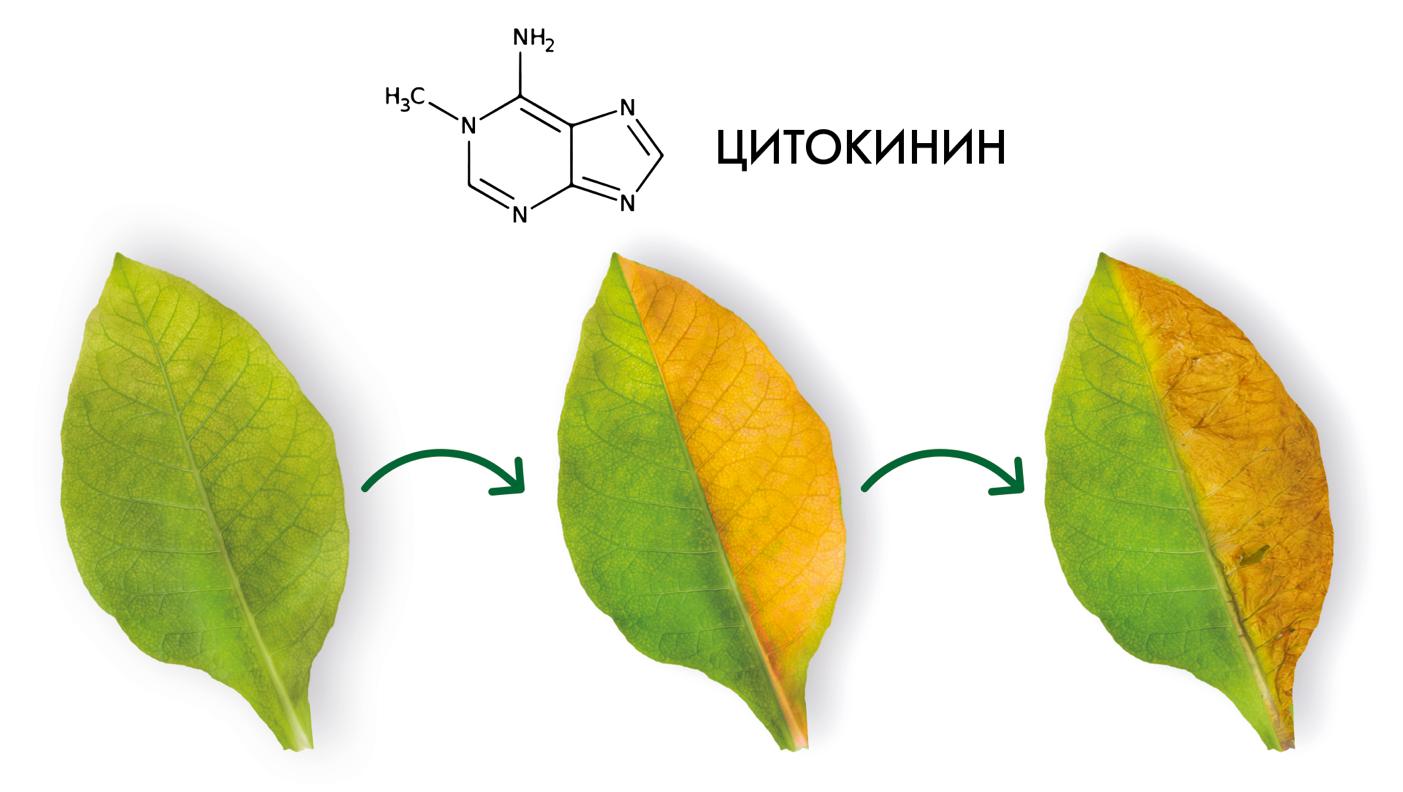
Рис. 14. Представлены листья махорки, где одну часть листа обработали цитокинином, что привело к остановке старения листа в виде пожелтения.
Функции цитокининового гормона молодости у растений?
Цитокинины синтезируются из азотистого основания аденина, одного из четырёх типов кирпичиков ДНК. Цитокинины запускают механизмы синтеза новой ДНК в клетке и контролируют S-фа́зу клеточного цикла у растительных клеток. S-фаза — фаза клеточного цикла, в которой происходит репликация ДНК. Стадия интерфазы, расположенная между G1 – G2 фазами. Главным событием S-фазы является репликация ДНК. Целью этого процесса является создание двух абсолютно идентичных хроматид. Клетка препятствует более чем одной репликации хромосомы, накладывая на ДНК в фазе G1 специальные пререпликационные комплексы, которые разбираются в S-фазе перед началом репликации. В почкующихся дрожжах белок Cdc6 разрушается, Orc2/6 фосфорилируется, и mcm-белки изгоняются из ядра, предотвращая новое прикрепление репликационного аппарата (ДНК-полимеразы) к ДНК после начала репликации.
Особое внимание акцентирую на то, что это его важнейшая особенность как гормона клеточного деления; причём в отличие от всех других гормонов (тоже обеспечивающих клеточные деления), позволяет ему усиливать не просто клеточное деление, но с учётом снятия, отмены программ как их онтостадийной продвинутости, так и морфологической дифференциации. Это подразумевает радикальную полную клеточную реювенилизацию и дедифференциацию. Только такого типа гормоны способны запустить механизмы регенерации не просто клеток, а тканей и органов.
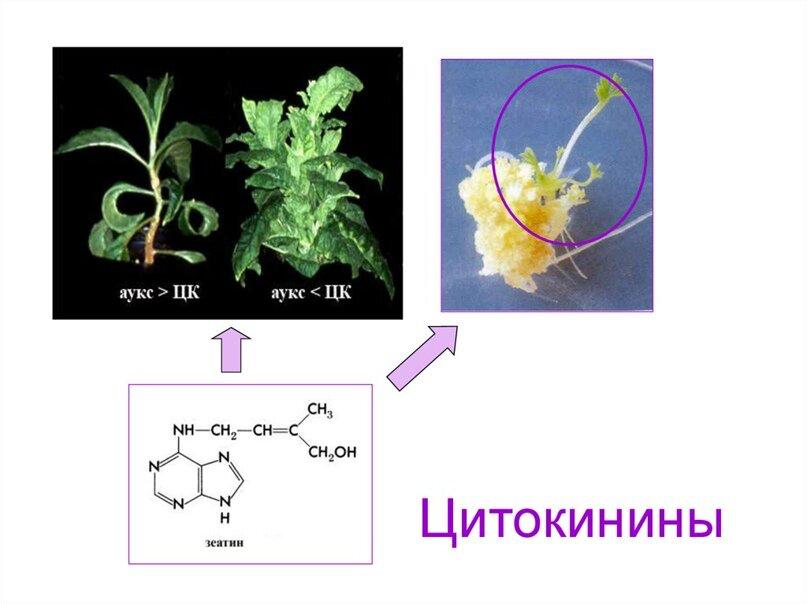
Рис. 15. На фото изображено физиологическое действие растительного гормона молодости цитокинина, где на первой картинке видно изображение растения, которое поменяло свои морфологические признаки онтостадийной зрелости на более ювенильные формы. Это пример стем-доминирования или радиксального доминирования, регулируемых преобладанием цитокининов над ауксинами. На второй картинке видно, что гормон цитокинин ведёт к закладке недифференцированной каллусной ткани (стволовых клеток обладающих высшей степенью потентности к регенерации). Эта бесформенная каллусная масса затем преобразовавлась в зачаток полноценного организма.
Очевидно, гормон может не просто дедифференцировать, но и стадийно реювенилизировать, что подразумевает аналогию отмены сенесцента. При этом закладка каллуса происходит в таком саморегулирующемся достаточном объёме, массе, чтобы из неё стал возможен запуск автоматически нового как эмбриогенеза, так и органогенеза с последующим образованием зародышевой ткани. Из зародыша затем развивается полноценный организм растения с корнями и листьями. В данном случае этот зародыш с тотипотентными возможностями может быть частично полноценным аналогом точки самовосстановления любой дифференцированной ткани в организме животного. Но только потентность здесь не превышает степень плюрипотентности, когда каждая ткань способна восстанавливать только сама себя. Очевидно с этим связано отсутствие как самого аналога цитокининового гормона в организме животных, так и отсутствие возможности полной регенерации. Возможно, взамен представлена система цитокинов, которые специфично представлены для каждого типа тканей.
В человеческом организме регенерация (закладка органогенеза заново) отсутствует практически полностью, но сохранён тканегенез. Более высокие уровни потентности глубоко заблокированы. Только лишь изредка встречается нечто подобное на каллусное перерождение у растений, например рубцовая ткань или в виде тератомных опухолей. Таким примером может быть тератома яичника – герминогенная опухоль яичника, содержащая нетипичные для данного органа ткани, являющиеся производными зародышевых листков. Может быть зрелой или незрелой.
Рис. 16. На рисунке изображена тератома. В 99% случаев новообразование является доброкачественным. Формируется в виде кисты, содержащей в себе эмбриональные ткани.
Рис. 17. Доброкачественная тератома развивается в виде новообразования с эластичной оболочкой. При вскрытии обнаруживают фрагменты волос, костей, зубов и кожи. Для доброкачественного новообразования характерно присутствие зрелых, дифференцированных тканей.
Следует отметить, что тератомы образуются только в эмбриональных тканях, что говорит о том что гормон её провоцирующий преобладает в эмбриональной ткани и присутствует в организме преимущественно в этот период, то есть может развиваться на эмбрионе или в яичниках. Во взрослом организме этих условий для присутствия соответствующего гормона нет.
Причём цитокинин настолько сильный гормон, что может способствовать закладке новых стволовых клеток и почек там где их не было, причем из паренхимы, то есть обычных покоящихся соматических клеток не способных в норме к этому, что говорит о регенеративных возможностях цитокинина. Это очень важно для случаев, когда апексы погибли или им не хватает мощи для отрастания. В этом случае нужен не рост, а регенерация. (Попутно отмечу, что в организме человека очень актуальна не только проблема реювенилизации, но отсутствия полной регенерации. Под термином реювенилизация понимается как отмену клеточного сенесцента, так и отмену старения ткани - онтофизис). Очень важно не путать эти два разных механизма, которые управляются и разными гормонами. Рост управляется ауксинами, а регенерация – цитокининами. Вместе они преодолевают старение. Можно подметить что ауксины – гормоны главных апексов, то есть верхних регуляторных структур, а цитокинины из нижних стволовых уровней, в данном случае из корней. Акцентирую на это внимание специально, чтобы подчеркнуть значимость ювенильных гормонов исходящих из низов. Очевидно, гормоны этого типа смогут преодолеть лимит Хейфлика. С ними не может быть связан гипоталамус и такие гормоны надо искать и у человека. Напомню, что у человека крайне не хватает или полностью отключена потенция на регенерацию, нет нужных для этого гормонов. Уточню, что ауксины способны вызвать прорастание почек, но не их закладку заново.
Попутно отмечу, что автором предложена концепция того, что лимит Хейфлика связан с отсутствием для этого специального внешнего доминирующего сигнала, который сопряжен с ювенилизацией. Попросту в организме животного на коммитативном уровне отключена возможность закладки достаточной мезенхимы, которая дополнительно инициирует ювенилизацию, то есть эта функция передана на внешние уровни регулировки. Поэтому в чистых культурах ткани эта доминанта подавлена и нужны усиливающие её условия.
Регулировка эмбриогенеза плода животных связана с превалированием нейрогуморальной над гормональной?
В свою очередь поиск ведущего гормона участвующего в эмбриогенезе плода животного показывает, что как такового его выявить не удаётся. Под влиянием каких гормонов происходит начальная фаза эмбриогенеза на стадии бластулы и гаструляции не известно. Но зато известно, что на этапе закладки органов эмбриона главным механизмом регуляции в этой системе становится нейрогуморальная. Обращает на себя внимание значимость инсулино-подобного фактора роста ИФР-1. Он участвует в эндокринной, аутокринной и паракринной регуляции процессов роста, развития и дифференцировки. Считается, что этот белок играет активную роль в процессах старения организма: мутации гена ИФР-1 приводили к увеличению продолжительности жизни у лабораторных животных.
ИФР-1 является важнейшим эндокринным посредником действия соматотропного гормона, поэтому называется также соматомедином С. ИФР-1 производится гепатоцитами печени в ответ на стимуляцию их соматотропиновых рецепторов. В периферических тканях именно ИФР-1 обеспечивает практически все физиологические эффекты соматотропного гормона. Его последствия были названы «неуправляемой инсулиноподобной деятельностью».
В принципе соматотропный гормон (СТГ) является гомологом гиббереллина – гормона роста у растений. Усиление его выработки в зрелом возрасте увеличивает акромегалию – диспропорцию скелета и изуродованию внешности. Посмотрите на рис. 2, где представлена рыба горбуша с гипертрофированной челюстью до такой степени, что это мешает ей нормально питаться и является причиной её мортуса. Поэтому поиск методов реювенилизации в этом направлении не даст результатов.
Факты подтверждающие концепцию значимости корней в усилении Жизненной Силы растения для увеличения продолжительности жизни данного вида
Приведу пример мощного пролонгирования продолжительности жизни другого хвойного дерева секвойи вечнозеленой, которая относится к числу самых долгоживущих деревьев на земле. Продолжительность её жизни достигает более двух тысяч лет. Но такую долговечность она может развить только в условиях богатых глубоких почв, где она развивает мощную корневую систему, которая и обеспечивает ей долговечность. Но если секвойю посадить на мелкую каменистую почву, то она быстро теряет свою долговечность, стареет через 20-40 лет, начинает суховершинить и отмирать. Здесь она не может развить достаточную мощную корневую систему, которая имеет развитую мезенхиму и может обеспечить дереву подачу непрерывного мощного сигнала на обновление и продолжение жизни. Примеры такого ускоренного старения секвойи показаны на фото ниже.

Рис. 18. Роща секвойи в Сочи в возрасте 60-лет, проявляющая все признаки суховершинения и преждевременного старения = сенилита = прогерия.

Рис. 19. Деревья секвойи на родине в Калифорнии в возрасте 500 – 1000 лет хорошо себя чувствуют и имеют нормально обветвлённую вершину. Доживаемость таких деревьев достигает несколько тысяч лет.
Корневое доминирование определяет Жизненную Силу кроны дерева
Здесь происходит децентрализация, когда значимость корней превалирует над значимостью кроны. Мощь корней отменяет программу на старение из кроны, откуда зарождаются сигналы на гиперапикальное доминирование и подавление сомы.
В человеческом организме отсутствует принцип радиксного (корневого) доминирования, что лишает его мощи репаративной мощи и неограниченного долгожительства.
В случае с человеком явно просматривается отсутствие принципа постоянно оживляющего радиксного доминирования, что является причиной нарастания гипергипоталамического доминирования (перезревание = матура) ведущего к постепенному не срабатыванию (подавлению) и угасанию стволовых клеток, а также подавлению и навязыванию сенесцентирования прогениторных клеток (аналог апексов 2-го, 3-го и т.д. порядков и брахиальных = адвентивных клеток растений). Накопление таких изменений также придаёт человеку все аналогичные признаки дряхлости организма. Только мощь зародышевых мезенхимных клеток может через их метаболом (сок) возродить прогениторы, а также все «точки самообновления» всех специфических тканей организма стареющего организма. Это означает, что если бы нам удалось поменять приоритеты с гипергипоталомического доминирования на усиление его антипода, противовеса, противодействующего его чрезмерному продвижению по онтостадийным ступенькам лестницы онтогенеза, за счет активирования зародышевых «точек самообновления» в виде мезенхимных и эмбриональных клеток, то мы бы смогли многократно раз отодвигать границы сенилита. В каждой ткани есть обязательно свои точки для непрерывного самообновления и если бы в этой ткани существовало равновесие, когда старые ткани постоянно обновляются на ювенильные (но с низкой степенью онтопродвинутости, то есть зрелости и не сенесцентирующие), то такая ткань могла бы существовать неограниченно. В данном случае тип роста апексных точек роста наружу (экзорост), который имеется у ветвей деревьев, заменяется на внутренний рост (эндорост), который имеется у животных, но с обязательным условием наличия механизма по выводу предшествующих клеток. В принципе это реальный механизм самообновления, но для этого нужны идеальные пейсмекерные уравновешивающие механизмы по апоптозу и выводу состарившихся линий клеток.
Принцип неотенического неограниченного существования
На противоположных принципах устроен онтогенез некоторых видов агав, которые продлевают своё существование за счёт образования постоянно новых вегетирующих столоновых отростков из спящих почек. По сути, это способ вегетативного размножения, который может продолжаться неограниченно. Именно этот тип почек обладает особенностью сохранения ювенильности и максимальной потентностью стволовых клеток. Такая коммитированная особенность позволяет агаве приобрести принцип неотенического неограниченного существования. Растение уходит в неограниченную неотению. Обеспечивает ей эту возможность мощный пул стволовых клеток = спящих почек. В данном случае можно говорить о принципе доминирования стволовых клеток второго порядка, стадийно более молодых, а не за счёт ведущей роли главных апексов, то есть более стадийно зрелых = апикального доминирования.
У молодых организмов человека имеется паритет между стадийно низкой зрелостью стволовых клеток гипоталамуса и стадийной зрелости стволовых клеток тканей. Их стадийная зрелость продвигается параллельно. Ювенильная сома на долго сдерживает зрелость гипоталамуса, но если в сому ввести факторы зрелости, то это даже у возраст-молодых организмов спровоцирует ускоренное старение гипоталамуса, а вслед за ним созревание и ускоренное старение всего организма. У старых организмов одновременно стадийно стары и стволовые клетки гипоталамуса и всех тканей.
Возможности цитокининового гормона в организме человека
Отмечу, что в организме животных он полностью отсутствует, тем не менее какое-то подобие его положительного действия можно обнаружить. Опишу эти случаи. Так, кинетин широко используют в косметике, замедляющей старение кожи, в том числе для уменьшения морщин, противодействия расширенными кровеносным сосудами, неравномерной пигментаций. Но это утверждают производители косметики, но не клинические исследования.
Один из таких продуктов – кинераза, омолаживающий крем и лосьон.
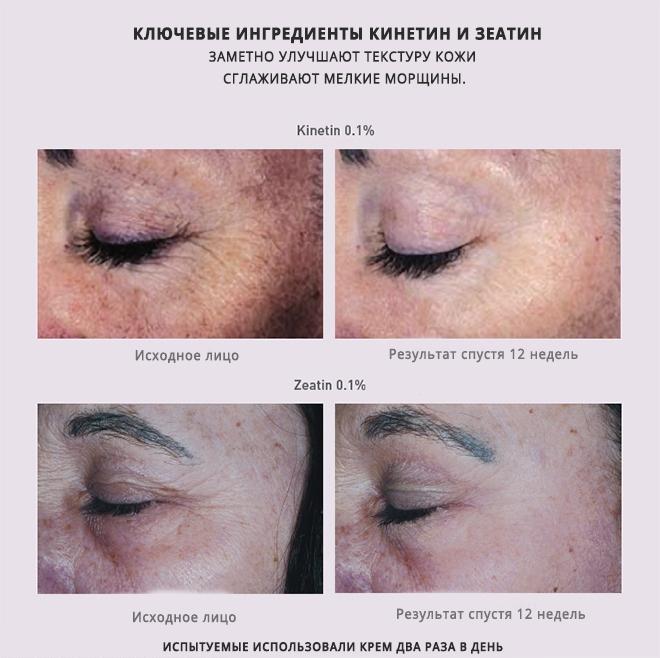
Рис. 20. На фото положительный пример применения цитокинина в косметических целях.
Поиск гормонов и веществ реювенилизаторов для организмов животных - аналогов цитокининовых гормонов растений
Выше были рассмотрены возможности цитокининового гормона растений для омоложения и снятия онтостадийного эффекта. У человека такой гормон не найден, но есть нечто похожее, что мало изучено. Важная деталь в том, что цитокинины можно получать путём автоклавирования и перегрева молок сельди – обильного источника ДНК. Перегрев вёл к распаду ДНК до новых молекул - фурфуриладенин. В последующем его назвали кинетин. Интересно, что кинетин снимает эффект апикального доминирования, вызванного ауксинами, и тем самым способствует закладке боковых почек, а не апикальных. Значит кинетин в роли контрпозита может противостоять эффекту гиперапикального доминирования = перезревания и тем самым делать растения более долгоживущими = многолетниками. Здесь следует увидеть аналогию между короткожителями - мышами и долгожителями - голыми землекопами. Один и тот же мезенхимальный ювенилизирующий эффект.
Точно также этот мезенхимальный эффект сказывается и у насекомых, например пчёл, где рабочие пчёлы живут 56 дней, а матки семьи от 3 до 7 лет. В данном случае пчёлы являются примером, когда объединены два процесса в одном: неотения и онтостадийное созревание, когда одно не мешает другому. Но здесь неотенические силы хотя и доминируют над онтостадийным продвижением, но не мешают ему, а не наоборот, как это часто встречается в природе. Поэтому неправильно утверждать, что неотения это всегда младенчество и отсутствие половозрелости. Очевидно, не следует ставить знак равенства неотении и фетальности, детству и эмбриональности.
Аналогичная ситуация как у пчёл наблюдается и у термитов.
Рис. 21. Морфологические разнообразие особей среди термитов, которое существенно влияет на длительность онтогенеза.
Существует ли аналогия между адениновыми радикалами цитокининов (гормонами молодости у растений) и митохондриальными РНК у человека?
На их аналогию наводят данные, что аденин, как один из нуклеотидов, из которых строятся нуклеиновые цепочки ДНК и РНК, является основой для цитокининовых молекул. В свою очередь некоторые митохондриальные РНК являются активаторами сенесцентных клеток, которые утратили способность к репликациям. Не означает ли это, что некоторые типы РНК могут играть роль цитокининовых гормонов? Возможно специализированные выделенные в отдельные структуры ткани с выработкой гормона ювенильности у человека недоразвиты? Тем не менее, этап ювенильности у человека длится до 12 лет и за счёт каких то механизмов он должен поддерживаться. Явно это связано с эпифизарным доминированием и его гормоном мелатонином. Возможно имеется связь с так называемыми мембранными пузырьками – экзосомами, наполненных разнообразными микрорегуляторными РНК (микроРНК). Это особые молекулы РНК очень небольшого размера, которые не отвечают за синтез тех или иных белков, а напрямую участвуют в экспрессии генов и способны управлять онтогенетическими программами. Это указывает на их гормоноподобное действие. Тем самым должны регулироваться уровень стволовых клеток, запускаться реювенилизация, а также преодолеваться клеточный лимит на репликации у сенесцентных клеток, им возвращается сила потентности, аналогичная камбиальной ткани. Таким образом, можно предполагать и вести поиск аналогичных клеточных универсальных гормонов, но работающих не на системном уровне организма, а на уровне омоложения всех типов клеток – универсальное омолаживающее средство. Такое гормоноподобное действие этих микроРНК и уникальная способность к реювенилизации и возобновлении репликаций сенесцентных клеток говорит об аналогичности их действия цитокининовым гормонам у растений. Но цитокинины вырабатываются на апексах корней, а микроРНК в «матрикальном апексе» = гипоталамусе, а также всех «точках самообновления». Такая централизация «гормона» объясняет очевидно и то, что в отличие от подлинных гормонов, микроРНК не обязательно попадать в кровь и разноситься ею по всему организму, вещество работает адресно в главном «матрикальном апексе», а через него на весь организм?! Первым стареет гипоталамус, а с ним параллельно и «точки самообновления».
Вызывает вопрос почему в организме животных аналог гормона ювенильности так и не найден в отличие от того, что он известен у растений и насекомых? Такая ситуация не означает, что его нет у животных вообще. В первую очередь это может быть связано с отсутствием принципа доминантности стволовых клеток (стем-доминирования), как это имеется у плюща и у сосны остистой. У большинства других видов родственных растений этот принцип отсутствует, это сокращает им частично потенциал жизни, укорачивает её, но они долго могут существовать не за счёт цитокининовой мощи, а за счёт ауксиновой, которая тоже стимулирует к росту, но относительно в более высокой онтостадийной степени продвинутости, с другим потенциалом потентности стволовых клеток. Только цитокинин даёт наиболее глубокий потенциал разблокировки потентности, вплоть до тоти- и плюрипотентности. В организме животных аналогичная ситуация. Подлинного глубокого = стадийного омоложения у них нет, то есть возврата до уровня эмбриональности у стволовых клеток, но присутствует репарация на уровне тканей с сохранением степени их онтостадийной зрелости. Эта зрелость «точек самообновления» у животных подобна сохранению онтофизиса у клоновых потомств, полученных от зрелых растений, которое проявляется в виде топофиза или циклофизиса [3]. Такого частичного самообновления иногда достаточно, чтобы существовать длительное время на высоких степенях онтостадийной зрелости. Онтостадийная зрелость сохраняет высокий уровень морфологической дифференцированности клеток, а также онтостадийного статуса. Но такая низкая степень разблокировки потентности не обеспечивает достаточный уровень для регенеративных возможностей.
Другой возможной причиной не нахождения гормона молодости у животных может быть то, что эта программа и вещество существуют только на эмбриональной фазе, то есть до родов плода. В это время организм вырабатывает огромное количество фетальных белков – фетопротеинов, после окончания беременности и родов количество фетопротеинов резко спускается почти до нуля. Постэмбриональные этапы жизни ребёнка и его ювенильности тогда определяются теми же принципами, что и регулировка ауксинами в организме растений, когда достижение высоких уровней его приводит к созреванию и переходу на новые зрелые стадии онтогенеза. Сами фетопротеины не являются гормонами, но определят статус. Очевидно их уровень определяют некие микроРНК, которые могут быть аналогами цитокининов.
Возможности молок сельди как концентрата ДНК – источника для образования адениновых дериватов, выступающих в роли индуктора и активатора точек самовосстановления, а значит реювенилизатора
Всё это заставляет вести поиск аналога растительного гормона цитокинина в организмах животных. В этом плане следует обратить внимание на источники адениновых веществ и разобраться в их механизмах действия. Рассмотрим подробнее оздоровительные возможности молок сельди как источника ДНК.
Сперматозоиды представляют собой гаплоидные клетки, то есть одинарный набор хромосом. Их дериваты после разрушения ферментами в желудке могут дать разнообразие адениновых комплексов, то есть веществ аналогичных гормонам и индукторам. В качестве рабочей концепции предполагается, что они в максимальной степени будут производить тотипотентные индукторы. Похоже что для этих целей приемлемо будет использовать рыбьи молоки. Результаты научных исследований показывают, что молоки сельди, кеты и др. содержат в составе высококачественную дезоксирибонуклеиновую кислоту = ДНК, которая распадается в организме на отдельные части из нуклеотидов. Высокая концентрация их может стать сигналом на усиление синтеза ДНК и запуск образования новых клеток, в том числе половых. Очевидно высокая концентрация аденина является сигналинговым веществом, которое «раскупоривает» ингибиторные статусы на подавление одной из фаз митозов. Этим можно объяснять сильный антивозрастной эффект при употреблении продукта. Как и цитокинин, аденин снизит уровень инактивации, повысит уровень потентности клеток, что позволит обеспечить запустить клеточную реювенилизацию и обойти лимит Хейфлика. Кстати, научный интерес представляет такой эксперимент в культуре ткани с повышенным содержанием аденина и других нуклеотидов в целях обойти лимит Хейфлика.
В поддержку этого являются и данные по применению молок лососевых в косметических целях. Для этого использовали ингредиент живого происхождения полинуклеатидов (ПДРН) – ДНК для сдерживания процессов хроностарения кожи, то есть омоложения кожи - от восстановления капиллярной сети до репрограммирования дермы.
Разработчики утверждают, что это «пролекарства», которые снабжают клетки необходимым количеством митогеннных дезоксирибонуклеотидов, дезоксирибонуклеозидов, а также азотистых оснований. По своей структуре ПДРН – это низкомолекулярные фрагменты, которые не вызывают генетической трансформации и являются абсолютно безопасными для организма человека. Выраженная эффективность ПДРН связана с их способностью замещать «сломанные» азотистые основания связывающие ДНК. Таким образом, препараты на основе полинуклеотидов не стимулируют работу старых функциональных структур в коже, а удаляют поврежденные участки клеток, способствуют их восстановлению и провоцируют рост молодых функциональных структур.
Иммунотропное действие ДНК из молок лососевых рыб
Показано, что это ДНК обладает сложным многогранным дозозависимым действием на иммунную систему организма, (наиболее активная доза -10 мг/кг веса мыши). Умеренно стимулирует клеточный и гуморальный иммунитет. ДНК модулирует продукцию цитокинов in vitro: исходно низкая продукция цитокинов усиливается, исходно высокая ослабляется. Введение ДНК из молок лососевых рыб обусловливает повышение уровня цитокинов, вырабатываемых преимущественно Тх1-клетками (ИФН -г), т.е. может способствовать развитию клеточного иммунного ответа. Действие ДНК на Т-систему иммунитета обеспечивает в эксперименте защиту макроорганизма от факультативных внутриклеточных патогенов (листерий).
Нуклеиновые кислоты в настоящее время широко используются в медицине в качестве иммуномодуляторов. Входящие в состав ДНК азотистые соединения (аденин и гуанин) являются структурной основой для низкомолекулярных биологически активных коферментов и кофакторов, лимитирующих биологические процессы во всех органах и тканях организма.
На основе этой нуклеиновой кислоты созданы фармакологические препараты — Деринат (ДНК из молок осетровых рыб), Полидан (смесь ДНК и РНК из молок осетровых рыб), Плацентекс-интегро (ДНК из молок форели), и биологически активные добавки (БАД) к пище — «Биостим» (ДНК из молок осетровых рыб), «ДНКаС», «ДНКаВИТ».
«Диэнай Карма» - препарат на основе олигонуклеотидов из рыбных молок – современная разработка для сдерживания старения за счёт активирования стволовых клеток
Принадлежит группе парафармацевтиков, является эффективной биологически активной добавкой, которая нормализует общее состояние человеческого организма, помогает укрепить иммунитет, восполнить свои природные жизненные силы и энергию. Обновляет внутренние структуры организма, заставляя регенерироваться ткани органов. Представляет собой иммобилизованные олигонуклеотиды (короткие фрагменты ДНК или РНК) и особый белковый комплекс. С помощью этого компонента клетки органов человеческого организма постепенно восстанавливаются, разрушенные ткани выстраиваются вновь, а их дальнейшее старение или деформация замедляется.
Восстанавливает Жизненную Силу клеток. Заставляет работать природную регенерацию организма и самообновление клеток «золотого фонда» организма – за счёт привлечения собственных стволовых клеток. Как итог, их дальнейшая деформация и старение будут замедлены. Биологически активная добавка угнетает фиброзные, диффузные и различные рубцовые изменение в паренхиме органов организма. Напомню что дистрофия и сенилит связаны со склеротизацией и фиброзными перерождениями дифференцированных клеток и сенесцентом стволовых клеток. Фиброз и кальциноз найдены даже в эпифизе. Действие препарата универсально, так как работает на клеточном уровне, не взирая на типы тканей и систем, а значит омолаживает все типы клеток и тканей. Очевидно, его действие связано с противодействием сенесценту любых стволовых клеток.
Важен для активного приёма при широком круге заболеваний, но, очевидно, в первую очередь при анизомалиях, то есть возраст-ассоциированных. Сочетается с основным лечащим препаратом, тем самым усиливая его свойства, и, также, в качестве профилактического средства. Заставляет усиленно работать регенеративные функции организма, укрепляет общий иммунитет.
Рекомендован для противодействия:
- опухолевые процессы доброкачественной и злокачественной природы;
- предотвращение процессов старения.
Эффект происходит за счёт обновления клеточного состава, включая стволовые клетки.
При этом показано, что препараты этой группы обладают противоопухолевым эффектом. А это согласуется с нашей концепцией, что возрастная тенденция предрасположенности к онкологизации - это всего лишь один из путей, вариантов проявления анизомалийных перестроек, связанных с сенесцентом клеток. Противодействуя сенесценту, мы одновременно противодействуем и другой их крайности - онкологизации. В тоже время это указывает на то, что возрастная гормонозависимая онкология не связана с мутациями, а есть одно из проявлений эпигенеза, анизомалией.
Как видим, действие препарата универсально, так как действует на все типы клеток сходно. Но, тем не менее, эффект даже если и имеет сходство с гормоном растений цитокинином, тем не менее, такого выраженного действия как у растений не имеет. Это цитокининоподобное действие, но не гормональное. Специфического гормона аналога в животном организме нет. Диэнай правильнее относить к клеточным нуклеотидным метаболитам, уровень концентрации которых определяет внутриклеточную детекцию готовности или достаточность сигналинговых веществ для запуска клеточных циклов. Очевидно, это генный медиатор-регулировщик циклов. Следовательно, он не является гормоном, как цитокинин, который вызывает каскад комплекса процессов, одним из проявлений которых является тоже накопление нуклеотидных метаболитов, запуск клеточных циклов и главное повышение уровня клеточной потентности, что означает одновременное снятие бремени высокой дифференцированности клеток, а также преодоление сенесцента = реювенилизация. Гормоны являются веществами-пускателями сложного комплекса процессов. Поэтому концентрация гормона многократно раз всегда меньше, чем уровень их метаболома, в том числе и нуклеометаболома, образующегося в результате их действия. Поэтому для достижения того же эффекта, что и у гормона цитокинина дозы олигонуклеотидов Диэная в данным случае должны быть многократно раз выше чем просто у гормона цитокинина.
Почему в организмах животных нет аналога гормона цитокинина – клеточного реювенилизатора?
Это связано с особой высокой степенью организации их ткани, где возврат на самые высокие уровни потентности (тотипотентность, плюрипотентность) недопустим. Такие возможности глубоким эшелонированием заблокированы, так как пробуждение этих потентностей может спровоцировать образование доброкачественных тератом, что несовместимо с возможностями существования организма животного. Образование клеток более низкого ранга дедифференцирования и открытия «ларчика» потентностей чревато гиперплазиями и доброкачественными опухолями.
Рис. 22. Во рту у 17-летнего мальчика обнаружили 232 зуба. Возможно это результат доброкачественной опухоли одонтома.
Поэтому гормон обеспечивающий полную реювенилизацию и регенерацию у животных не предусмотрен. Тем не менее, использование метаболитных нуклеотидых регуляторов для снятия сенесцента вполне целесообразно и безопасно. До уровня тоти- и плюрипотентности они не опускают.
Фактор Яманаки
О возможности омоложения человеческих клеток
Открыты химические вещества, позволяющие индуцировать перепрограммирование, то есть обратить вспять старение и восстановить функции клеток. Выяснено что экспрессия специфических генов, называемых факторами Яманаки, позволяет превращать взрослые клетки в индуцированные плюрипотентные стволовые клетки (иСК). Для этого определены шесть химических коктейлей, которые восстанавливают эти белки и геномные профили транскриптов до молодого состояния и обращают транскриптомный возраст вспять менее чем за неделю.

Рис. 23. В эксперименте участвовала быстро стареющая линия мышей. Возраст мыши (справа) был увеличен на 150% по сравнению с её братом (слева) путём нарушения эпигенома.
Исследования зрительного нерва, мозговой ткани, почек и мышц показали многообещающие результаты: улучшение зрения и увеличение продолжительности жизни уже наблюдалось у мышей, а недавно появилось сообщение об улучшении зрения и у обезьян.
Получены положительные результаты по обращению слепоты у обезьян. Таким образом, было доказано, что старение можно не только сдерживать, но и обращать вспять. Открытие даёт возможность обратить вспять процесс старения с помощью одной таблетки, что может найти широкое применение, начиная от улучшения зрения и заканчивая эффективным лечением многочисленных возрастных заболеваний.
Информация в клетках запрятана как на генетическом, так и на эпигенетическом уровне. Сенесцент клеток обусловлен эпигенетически. Одним из признаков старения организма на клеточном уровне считается потеря этой эпигенетической информации. Клеточное старение приводит к остановке клеточного цикла, потере клеточной идентичности, а затем и полной дисфункции клеток. Для организма это выражается в снижении способности к заживлению ран, ремоделированию тканей, предотвращению рака за счет остановки деления старых и поврежденных клеток, а также в развитии других типичных возрастных заболеваний.
С помощью веществ фактора Яманаки можно перезапустить программы, превращая взрослые и даже старые клетки обратно в стволовые. Другими словами происходит сбрас часов клетки на ноль, возвращая взрослые клетки в их эмбриональное состояние. Но некоторые авторы трактуют это по другому и утверждают, что процесс не влияет на клеточную идентичность, то есть старая клетка кожи после процедуры не становится стволовой, а остается клеткой кожи, только несколько более молодой. Метод назвали EPOCH - «эпигенетическое программирование здоровья старых клеток».
Появилась надежда, что эти клетки, лишенные характерных признаков старения, можно будет использовать для восстановления и омоложения поврежденных органов. Например, более молодые и здоровые нервные клетки могут заменить клетки мозга, погибшие в результате инсульта, или клетки кожи, повышающие выработку коллагена, можно вводить непосредственно в незаживающие раны.
Но оказалось есть и проблема: факторы Яманака слишком сильно перепрограммируют клетки. Клетка, возраст которой равен нулю дней, не может посылать электрический нервный сигнал или вырабатывать коллаген, ни выполнять какие-либо другие функции. Как и стволовая клетка, это не более чем сгусток возможностей.
Факторы Яманаки как средство омолаживающего действия через эпигенетические механизмы
Доказано, что взрослые клетки можно перепрограммировать путём пересадки их ядра в яйцеклетку. Исследования Ш. Яманака углубили эти доказательства: он показал, что экспрессия четырех белков трансформировала соматические клетки мыши в клетки близкие к эмбриональным стволовым – в так называемые индуцированные плюрипотентные стволовые клетки = iPSCs.
Эти данные обнадёжили весь научный мир. Но при этом ученые не учитывают, что сенесцентность клеток это не самостоятельный процесс, идущий снизу на уровне клеток, а является следствием онтофизисного эпигенеза. Для создания омоложенных тканей и выращивания органов метод весьма привлекателен; но на уровне целостного организма он не позволит решить проблему, так как его будут нивелировать процессы более высокого порядков, в том числе и сенелит, в частности слабая мощь поддержки ювенилизирующих гормонов со стороны эпифиза, но зато мощное давление гормонов провоспаления = подавления.
До сих пор существуют препятствия в получении индуцированных плюрипотентных стволовых клеток, преодоление которых поможет значительно повысить эффективность и скорость генерации iPS-клеток.
К тому же процесс получения индуцированных плюрипотентных стволовых клеток крайне неэффективен. Полное перепрограммирование соматических клеток в iPSCs может занять целый месяц, при этом только одна из тысячи клеток станет плюрипотентной. Более того, результаты целого ряда исследований свидетельствуют о том, что при всей своей пластичности iPSCs не являются абсолютным эквивалентом эмбриональных стволовых клеток.
Установлено, что через 48 часов эти факторы, как правило, связываются с регуляторными элементами генов, так называемыми энхансерами, находящимися далеко от регулируемых ими генов, а не с самими генами-мишенями. Это говорит о том, что эти факторы являются «пионерными», открывающими закрытые хроматиновые структуры на самой ДНК и облегчающими процесс перепрограммирования клеток, делая фрагменты-мишени генома доступными для считывания.
Обнаружены большие области генома, рефракторные к связыванию с факторами перепрограммирования в течение первых 48 часов, которые, однако, в конечном итоге активируются и являются необходимыми для образования индуцированных плюрипотентных стволовых клеток. По сути дела, существуют большие фрагменты человеческого генома физически сопротивляются связыванию с этими факторами. Поэтому их надо уметь блокировать. Блокирование ферментов, катализирующих синтез этой модификации, значительно ускоряет процесс перепрограммирования клеток.
Возможности пыльцы растений в качестве источника нуклеотидов для сохранения молодости
Выше мы обсуждали возможности рыбных молок как источника нуклеотидов и адениновых веществ в качестве аналогов процитокининового гормонального действия. В данном случае эти молоки являются гаметами, клетками для оплодотворения яйцеклетки. Но такие же гаметы имеются в изобилии и в растительном мире – это пыльца растений. Рассмотрим их возможный потенциал физиологического действия в плане клеточной реювенилизации, в том числе и против их сенесцента.
Пыльца содержит элементы, которые могут улучшить стабильность ДНК, нейтрализовать свободные радикалы и свести к минимуму их воздействие на организм.
Клетки нашего организма, питаемые антиоксидантными ферментами (а в пыльце 104 фермента), оказывают положительное влияние на замедление старения клеток.
Это способность организма поддерживать здоровые и жизненно важные ткани, а также замедлять старение и, следовательно, увеличивать продолжительность жизни. Предполагается, что это связано со снижением окислительного стресса с помощью пока в основном нераскрытых механизмов, и эти изменения в окислительно-восстановительном статусе, по-видимому, являются одним из основных механизмов продления жизни и благотворного воздействия на здоровье.
Экстракты пыльцы сосны могут замедлять старение клеток.
Пыльца увеличивает количество полезных антиоксидантов, таких как глутатион и СОД или супероксиддисмутаза.
Глутатион необходим для метаболизма некоторых витаминов и минералов, таких как железо, витамин С, витамин E и участвует в транспорте аминокислот. Глутатион также подавляет рост раковых клеток, нейтрализует свободные радикалы и важен для синтеза и производства ДНК.
Кроме того, глутатион содержит фермент глутатион-S-трансферазу, который является ферментом, выводящим токсины из печени.
Еще одно преимущество пыльцы заключается в том, что она поддерживает здоровое производство СОД, которое обычно имеет тенденцию к снижению по мере того, как мы становимся старше.
СОД является питательным веществом, препятствующим старению, и естественным антиоксидантным ферментом, вырабатываемым в организме, и играет важную роль в качестве поглотителя свободных радикалов, который также защищает от повреждения ДНК.
Пыльца является отличным средством для профилактики возрастных заболеваний у людей.
Примечательно, что пыльцу рекомендуют при следующих возрастных анизомалиях:
- ПМС,
- гиперплазия простаты,
- хроническая усталость, депрессия,
- заболевания печени,
- сердечно-сосудистая система,
- гематологические проблемы,
- преждевременное одряхление,
- нервной системы, астения,
- иммунитета.
Способ применения пыльцы.
Утром, в обед, до еды за 30 минут, или вечером, чтобы уменьшить чувство голода. Суточная доза – 1-2 чайные ложки. Можно также добавлять пыльцу в смузи и коктейли. Наибольшую пользу даст способ тщательного рассасывания её во рту до полного растворения.
Дневная рекомендуемая доза пыльцы должна не превышать 12 г, то есть до 4-х чайных ложек в день. Но это уже будет покрытие полной дневной потребности в метионине, поэтому надо учитывать, что часть его еще будет поставляться и с другой пищей. Поэтому лучше ориентироваться на 1-2 ч. л. Это связано с тем, что в этом количестве содержится максимально допустимая суточная доза - 12 миллиграмм метионина. Дальше передозировка. При тяжелых болезнях дозы можно повышать. Детям превышение от детских доз не вредит.
Более высокое суточное потребление может применяться для физически активных людей, спортсменов или тех, у кого высокий дефицит питательных веществ.
Парадокс метионина
Ряд источников обращают внимание на рекомендации ограничения применения пыльцы. Это связано с тем, что содержит аминокислоту метионин, действует как антиоксидант, который должен предотвращать процесс старения и омолаживать клетки тела. Кроме того, улучшает качество жизни при старении и ослабляет сопутствующие заболевания, связанные со здоровьем, такие как ожирение, сахарный диабет и когнитивные расстройства. Однако, ряд исследований на мухах дрозофилах, мышах показывают обратное – ограниченное потребление продуктов, богатых метионином, способствуют продлению жизни. Как же так: с одной стороны он омолаживает, а с другой – состаривает? Ряд авторов утверждает, что противоположный эффект начинается при завышенных дозах. Но это не верно, парадоксальный эффект начинается только с возрастом.
Почему такой парадоксальный эффект?
Метионин широко используют для лечения и профилактики таких возраст-зависимых заболеваний, как: болезнь Паркинсона, Альцгеймера, диабет, цирроз, мастопатии.
Метионин рекомендуют: от раннего старения кожи, при ломкости ногтей, атеросклерозе, рассеянном склерозе, алкоголизме, хронической усталости и мн. др.
О возможных механизмах связи метионина и старения – одна из версий.
Очевидно, ограничение потребления пищи может восприниматься организмом, как угроза жизни от голода. Голодание – это вероятность снижения плодовитости, а значит голодание — это прямая угроза всему виду. Организм на такую возможную угрозу реагирует увеличением продолжительности жизни и удлинением его репродуктивной функции. Тем не менее, это не дает окончательного ответа и противоречит другим примерам из природы. Так пересадка деревьев лиственницы в каменистую бедную почву привело наоборот к преждевременному заложению шишек, то есть ускорило генеративное созревание и плодоношение, ускоряется созревание и старение организма. Точно такое же произошло с деревьями секвойи, когда в молодом 60-летнем возрасте начала суховершинить, стареть и отмирать. То есть плохое питание в молодости у некоторых видов растений вызывает ускорение генеративной зрелости. Это можно объяснить плохим развитием их корневой системы, а значит недовыработкой цитокининов с преобладанием значимости верхних апексов, что и ускоряет закладку шишек, укорачивает вегетативную ювенильную фазу жизни организма. В человеческих организмах регулировка зрелости происходит иначе. Так, дети, которые питаются неполноценно (недоедание, недостаточное количество питательных веществ) могут испытывать задержку полового развития, в отличие от сверстников, которые питаются полноценно. Это продлевает ювенильную фазу существования организма, что чаще совпадает с продлением онтогенеза. Ограничение калорийности пищи или приёма метионина и цистеина (серосодержащие аминокислоты) в зрелом возрасте тоже продлевает продолжительность жизни мышей на 30-50%.
Похоже, что организм вычисляет объём потреблённой пищи не по съеденным калориям, а по количеству метионина в организме. Кроме того, есть мнение о том, что именно по количеству метионина организм вычисляет и количество других аминокислот, необходимых для образования белков.
Метионин активизирует киназу mTOR, которая отвечает за регулировку поддержания клеточного гомеостаза, развития опухоли и старения.
Этот белок занимает одно из центральных мест в процессах старения живых организмов, стимулируя развитие возрастных патологий, укорачивающих жизнь. Серин / треониновая протеинкиназа mTOR регулирует поддержание клеточного гомеостаза путём координации, сопряжения ряда процессов, включая транскрипцию, трансляциию, метаболизм и аутофагию с доступностью аминокислот, факторов роста, АТФ и кислорода. Киназа mTOR является компонентом двух белковых комплексов, mTORC1 и mTORC2, которые различаются по своему составу и регулируют разные клеточные процессы. Неконтролируемая активация киназы mTOR наблюдается в клетках большинства опухолей, а таже при диабете, нейродегенеративных и некоторых других заболеваниях. Вырисовывается картина, что mTOR подавляет аутофагию путём утилизации белков, но на фоне усиления их синтеза.
Другой возможный фактор того, что метионин способствует быстрому старению.
Киназа mTOR – это внутриклеточный протеин, который работает как ключ-вектор, триггерный переключатель метаболических процессов, перенаправляя их потоки по заданному направлению с учетом многочисленных других факторов. От её активности зависит очень многое, включая и продолжительность жизни. Всё дело в том, что активизированная киназа mTOR сокращает продолжительность жизни из-за того, что при таком возбуждении включаются механизмы анаболизма, как в молодости, когда вырабатываются всё новые и новые белки, происходит неуправляемый «белковый хаос», диссонанс потребности и реальной поставки, что приводит к тому, что старые белки не успевают полностью утилизироваться. В молодости существует гормональный баланс, когда этот анаболизм нужен, что обеспечивает рост (аналог вегетирвания), что даёт возможность легко их утилизировать и перенаправить по назначению. Переизбыток метионина здесь только на пользу, так как способствует как механизмам роста, так и поддержке клеточной реювенилизации. Но с возрастом и гипоталамическими регуляторными перестройками происходит переформатирование и на уровне клеток в виде онтофизиса, когда происходит перестройка их метаболизма и переход на новые программы и принципы существования. Клетки ориентированы больше на катаболические процессы, с последующей динамикой уровня их зрелости происходит повышение их резистентности к системам сигналинга. Они становятся менее чувствительными, более «глухими», а значит менее управляемыми и функциональными. А это пролог к сенесценту. Следовательно, здесь на клеточном уровне закладываются основы для старения всего организма.
При этом все эти возрастные перестройки проявляются в виде эпигенетических надстроек и передаются в поколениях дочерним клеткам. Это связано с тем, что дочерние клетки образуются из стволовых клеток не самых высоких уровней потентности типа плюрипотентность, а в основном из прогениторного типа и не способны "скинуть приобретённый груз". Итак, у зрелых клеток от взрослых организмов происходит переход на новые программы и принципы регулировок, что обусловлено коммитативными особенностями клеток животных. К этому всё предначертано.
В результате у зрелых клеток избыток метионина приводит к дисбалансу белков, который должен быть компенсирован автоматически механизмами аутофагии, но этого нет, так как её блокирует mTOR. Очевидно такая патологическая реакция на избыток метионина у взрослых организмов является нормальной, а механизмы противодействия ей не предусмотрены. Возможно это эволюционный механизм запуска управляемого по времени программ сенилита и мортальности. Но, возможно, здесь скрываются другие противоречия, которые могут вскрыться при противодействии им.
Вместе с этой блокировкой аутофагии и накоплением специфических белков запускаются автоматически механизмы изменения метилирования хроматина. Меняется электростатическая натяжка хромосом со всеми многочисленными последствиями, в том числе и укорочение теломер из-за перекрытия работы теломеразы. В молодости до фазы зрелости организма, до перехода на новый гормональный фон теломераза работает полноценно.
Похоже нутриентные стимуляторы метаболизма и обновления клеток (метионин и др.) при их избыточности необходимо обязательно сочетать с ускорением аутофагических процессов, которые можно обеспечить только мягким голоданием или интервальным 1-разовым питанием.
Проще говоря, если киназа mTOR активизирована, то это ведёт к тому, что не утилизированный белковый «мусор» засоряет весь организм, отчего организм ускоренно стареет.
Становится фактом, что метионин и ещё некоторые аминокислоты как раз и активизируют, возбуждают белок киназу mTOR.
Метионин при его избытке преобразовывается в S-аденозилметионин и является донором метильных групп, которые участвуют в избыточном метилировании нуклеиновых кислот - основы эпигенетических перестроек.
Эпигенез влияет на сенесцент клеток. Но это возможно только при определенных условиях. На молодые и растущие организмы это не сказывается.
Биологический возраст или эпигенетические часы как проявление онтофизиса.
Очевидно, этому способствует онтофизис, то есть отпечатывание возрастных изменений на клеточном, а точнее эпигенетическом уровне. Но этот эпигенетический уровень коррелирует с онтофизисными перестройками на уровне всего организма, начальный ключ для всех опосредуется через драйвер эпифиза. На каждом этапе онтогенеза, начиная от эмбрионального и кончая глубокой старостью, когда начинается этап превалирования сенесцентных клеток, имеются свои специфические для них онтофозисные фенотипы, паттерны, включая и паттерны старения. На уровне клеток онтофизис обозначается через череду меток на эпигеноме на длинной линии жизни. Задают, «рисуют» эти метки паттерны, которые обозначает эпифиз через гипоталамус и другие оси. У каждого вида животных имеется своя предписанная длина линии жизни = онтодианность*. Онтодианность зависит от возможности самореализации эпифиза и его сонастройкой с системой оси гипоталамуса. Эта сонастройка опредляет принципы длительности всех основных этапов на онтодианной линии как:
- квазиэпифизарного доминирования (фетальный период эмбриона),
- эпифизарного доминирования,
- паритета эпифиз-гипоталамус,
- гипоталамического доминирования,
- гипергипоталамического доминирования,
- гипоталамическая инволюция.
Подмечу что при рождении ребёнка он находится всё время в состоянии сна, а значит нет работы механизма циркадианности* эпифиза, а только персистирования. Поэтому мною этот период обозначен как квазиэпифизарного доминирования*.
Любые жизненные изменения: болезни, возрастные перестройки - откладывают свой отпечаток на эпигеноме. По видимому, эти отпечатки в виде интенсивности метилирования в наибольшей степени проявляются там, где наиболее активно функционируют участки хромосом. Чтобы открыть-экспрессировать одну группу генов необходимо при этом закрыть другую группу контрпозитных генов. Поэтому одни части хромосомы метилируются сильнее, а на другой - идёт ослабление метилирования (репрессия – экспрессия).
Концепты эпигенетических часов и реостата хроноса.
Все эти метки являются переходными этапами эпигенеза. Это позволило Стиву Хорвату выдвинуть концепт эпигенетических часов. Из этого исходят и понятия биологический возраст и эпигенетический возраст. Последний зависит не только от внутренних программ развития, но и от внешних факторов среды, когда он может как обгонять, так и отставать от первого. Задача научиться раздвигать разницу между этими возрастами. Но этот концепт не совсем точно отображает суть вопроса, это не столько часы сколько проявление степени продвинутости по ступенькам онтогенеза. Как таковых нет эпигенетических часов, а есть механизм продвижения по некому реостату реализации определенных программ и механизмов, эта реализация идёт с определённой последовательностью и скоростью. Это более правильно определить принципом как реостат последовательной хронологической реализации на онтодианной линии онтофизисных фенотипов (внешне проявленных) и паттернов (внутренних, скрытых параметров), то есть последовательности возможных сценариев развития. Проще реостат хроноса*.

Рис. 24. Принцип действия реостата подобен принципу действия онтофизиса при продвижении его вдоль линии жизни – спирали из последовательных витков обмоток (онтогенез), на каждом участке которой имеется свой виток, то есть комплекс генопрограмм, онтопрограмм реализации геногенеза.
На рис. 24 изображен принцип действия реостата, который имеет сходство с принципом реализации онтогенеза, где на каждом этапе продвижения имеется фиксация, коммитирование* своего онтофизиса. При продвижении реостата жизни, то есть продвижении вдоль линии жизни, которая представляет собой подобие череды последовательных витков обмоток (спирали = онтогенеза) происходит смена онтостадий. На каждом участке спирали-онтогенеза имеется свой виток, то есть комплекс генопрограмм, онтопрограмм реализации геногенеза.
Давайте повнимательнее посмотрим, как работает реостат. Его принцип основан на том, что мы можем менять сопротивление провода, следовательно можем регулировать силу тока и напряжение в электрических цепях. Согласно закону Ома U=IR. Напряжение равно произведению силы тока на сопротивление, то есть резистентность. Длину керамической трубки можно представить как онтодианную линию, а количество пройденных витков – как реализация паттернов и фенотипов онтофизиса, которые если их сгруппировать являются проявлениями онтоэтапов. Количество пройденных витков – это сопротивление, резистентность. С возрастом эпифиз и гипоталамус становятся по очереди всё более резистентными, нечувствительными, чему способствует, очевидно, как накопление сенесцентности на уровне клеток, так и сами онтофизисные перестройки. Это аналогия увеличения резистентности на реостате со степенью увеличения отодвижения ползунка по реостату. Как уменьшить сопротивление = R в этой формуле – уменьшить передвижение драйвера-ползунка по виткам онтофизиса. Ток = I в этой формуле можно понимать как Жизненная Сила, а напряжение = U как количество пройденных витков на онтодианной линии. Чем меньше пройденных витков, тем выше ток и меньше сопротивление, а значит больше степень фенотипов молодости. Следовательно, вся проблема в ползунке = эпифизе.
Предположим, что длина керамической трубки с намотанными на ней витками представляет собой заданный потенциал жизни, потенциал возможностей, а длина этой трубки отвечает за онтодианность. Допустим длина её 120 см, а это даёт возможность прожить до 120 лет. Это означает, что на пути этой онтодианнй линии с увеличением прожитых лет увеличивается резистентность, сопртивляемость Жизненной Силе, сижается чувствительность управляющих-оптимизирующих всё органов. На конце такой трубки = онтодианной линии сопротивление доходит до критически больших величин, так как коридор гормезиса резко сужается, а Жизненная Сила = напряжение – до критически малых, то есть доходит до границы, за пределами которой несовместима жизнедеятельность и это является началом неизбежного - фазы мортуса, кототрый реализуется за счёт активных механизмов феноптоза. Следует отметить, что мортус или сброс, обрыв это не случайностный, то есть не стахостический процесс, а активный, так как организм на последних этапах давно находится за границами гормезиса, оптимума, большинство систем работает вразнос, а значит крайне активизированы силы анизомалий* (возраст-зависимые заболевания).
Основными механизмами реализации феноптоза являются анизомалии. Они не случайны, а не избежны. Они не являются проявлениями внешних патологических факторов, то есть не патогенны, а сугубо эндогенны, являются продолжением эндогенного развития за пределами гормезиса, когда включаются механизмы на самоликвидацию. Это аналогично тому, как включаются механизмы апоптоза в осенний сезон и все листья на дереве до единого сбрасываются, исключений нет. Происходит это за счет гормона старения – абсцизовой кислоты; листья не осыпаются сами, допустим из-за заморозков, а их активно к этому сбросу подводят. Точно также этот же гормон абсцизовая кислота является причиной феноптоза у растений, то есть причиной их короткожительства, например это у сельдерея, у которого феноптоз запускается на вторй год жизни и связан с доминированием в онтогенезе абсцизовой кислоты, вырабатываемой в цветочных зонтиках и семенах. Такой же процесс ограничения онтогенеза имеется и у агав в период их зацветания на восьмой год. Это эндодианный процесс феноптоза. У деревьев идёт активный листопад, а у человека с наступлением сенилита идёт активный феноптоз, в возрасте 75-100 лет идёт активный «сброс» всей возрастной популяционной группы. Ведущими анизомалиями являются проблемы в сердечно-сосудистой системе и онкология, преимущественно гормонозависимая. Вслед за ними идут анизомалии в системах почек, костной системе – остеохондроз, системе крови, печени…
Апоптоз листьев у деревьев можно притормозить за счёт обработки цитокинином, а у человека, очевидно, за счёт мелатонина и репарации в эпифизе. Но проблема в том, что те дозы мелатонина, которые работали на этапе эпифизарной молодости, уже не сработают при применении их на других этапах. Дело в том, что за это время продвинулось созревание оси гипоталамуса, прошла его элевация и, возможно и инволюция, а значит он уже не чувствителен к прежним дозам мелатонина, которые его тормозили ранее. В этом случае, чтобы «пробить» эту «заглушку» требуются значительно другие дозы, многократно, десятки раз выше тех, что были в период юности. Возможно, это уже будут токсические дозы? Возможно, для репарации в этой системе нужно будет отработать механизмы аутофагии, сенесцентолитиков, то есть освобождение от старых гликолитических клеток за счёт апоптоза, а также циклические курсы по перезапуску эпифизарной оси на персистирующий режим. Это нечто подобное тому, когда у растения запускают механизмы этиоляции (временный рост в темноте), в условиях которой они перезакладывают не онтофизисные или генеративные точки роста, а ювенильные и вегетативные. Онтофизисные, то есть зрелые можно сравнить с сенесцентными. Но у человека этой перезакладке мешают механизмы циркадианности, которые необходимо научиться отменять.
Гормон стресса и гиперзрелости у растений как фактор их апоптоза и феноптоза – аналог провоспалительным веществам у человека как фактора реализации механизмов апоптоза и феноптоза через анизомалии.
Напомню, что у ряда растений ограничение онтогенеза происходит за счёт гормона старения – абсцизовой кислоты; листья не осыпаются сами, допустим из-за заморозков, а их активно к этому сбросу подводят эндогенные программы. Точно также этот же гормон абсцизовая кислота является причиной не только листопада, то есть апоптоза у отдельных органов, но и феноптоза всего растения, то есть причиной их короткожительства. Примером этого является сельдерей, у которого феноптоз запускается на вторй год жизни и связан с доминированием в онтогенезе абсцизовой кислоты, вырабатываемой в цветочных зонтиках и семенах. Смотрите рис. 7. Такой же процесс ограничения онтогенеза имеется и у агав в период их зацветания на восьмой год, рис. 32. Это эндодианный процесс как отмирания листьев в активный листопад, так и феноптоза всего организма.
Точно также у человека с наступлением сенилита идёт активный феноптоз, когда в возрасте 75-100 лет происходит активный «сброс» всей возрастной популяционной группы. Запускают активно этот процесс группа провоспалительных гормонов цитокинов, в частности, например интерлейкин-17 (IL -17), который найден в обилии в стареющей коже у стариков. Именно он играет решающую роль в генезе старения кожи. Посмотрите рис. 12 и 13. Именно эти провоспалительные цитокины являются причной всех анизомалий. Через анизомалии запускается апоптоз. Старение, например кожи или сосудов, не происходит само по себе, а инициируется активно. Кстати, известна антиинтерлейкиновая терапия по омоложению кожи. Определено, что некоторые иммунные клетки в коже экспрессируют высокие уровни IL-17. Этот же интерлейкин ответсвенен и за аутоиммунное заболевание как псориаз, так и эндогенные процессы связанные со старением. Найдена связь между иммунными клетками, воспалением и старением. Кожа является местом обитания для подвижных иммунных клеток, которые играют решающую роль в эндогенных и экзогенных процессах кожи в том числе и старении.
С возрастом присутствие некоторых из этих иммунных клеток, а именно гамма-дельта Т-клеток, врожденных лимфоидных клеток и CD4+ Т-клеток, значительно увеличивается в коже. Эти же клетки также начинают экспрессировать очень высокие уровни провоспалительного цитокина IL-17.
Точно также, как старение кожи является одной из анизомалий сенилита, так и всё остальное множество анизомалий обязательно сцеплено с неминуемыми провопалениями в других органах. Именно провоспаление является первичным условием для провоцирования и поддержания анизомалий. Анизомалии и являются механизмами реализации феноптоза.
Также мелатонин демонстрирует отчетливые антистрессорные свойства.
Уровень мелатонина определяет уровни провоспалительных гормонов.
Показано, что чем больше в организме уровни мелатонина, тем ниже уровни провоспалительных цитокинов, и наоборот – чем больше цитокинов, тем меньше мелатонина.
Имеется ряд клинических и экспериментальных доказательств влияния мелатонина на различные патологические состояния, сопряженные с воспалением. Среди них воспалительные поражения суставов, внутренних органов (типа язвенного колита), периферических нервов. Мелатонин является синергистом для НПВС (неспецифические противовоспалительные вещества).
Представлены также многочисленные сведения о способности мелатонина (3—6 мг) ослаблять головную боль разного происхождения. У мелатонина есть отчетливые обезболивающие свойства, которые должны быть непременно востребованы при болевых синдромах разного генеза.
Авторская классификация онтоэтапов.
Ранее в биологии было известно несколько классификаций, но автором предложена иная классификация, которая построена на принципах переходных изменений высших регулировочных систем, а точнее последовательность, очередность доминантности тех или иных структур.
Классификация онтоэтапов по Г. Гарбузову:
- Эмбриональный период - регулируется фетальными гормонами.
- Квазиэпифизарного доминирования – постнатальный период пока ребёнок находится преимущественно в состоянии сна до 1-го - 2-х лет.
- Эпифизарного доминирования – период с 2 до 12 лет и начала пубертата, то есть эпифизарной молодости, неполовозрелости.
- Паритета эпифиз-гипоталамус – с 12 до 21 года – этап юности.
- Гипоталамического доминирования – этап зрелости, адальтуса.
- Гипергипоталамического доминирования – этап климакса и перезрелости, матуры.
- Гипоталамическая инволюция – этап сенелита или геронтоса.
- Стем доминирования* – этап мортальных болезней и мортуса = сброса = феноптоза.
Скользящий ползунок является эпифизарным драйвером. Только если в реостате ползунком управляет человек из вне, то в оси эпифиза его переключения осуществляются автоматически по мере достижения определённых критических параметров. Отключает эпифиз по мере достижения созревания гипоталамус, он является перекрывающим эпифиз клапаном, заглушкой. Точнее не сам гипоталамус, а вся ось, которая ему подчиняется. Таким образом, онтогенез является чередой онтодианных отрезков. Поскольку эпифиз — это драйвер, он постоянно находится в «движении», самоизменении, и при этом заводит в движение, изменения ему подчиняемые системы. Каждый этап онтогенеза сопровождается специфичными для него изменениями в оси подчиняемой эпифизарному драйверу. Точнее это гипоталамо-эпифизарная ось.
Принцип действия эпифизарной оси.
Её можно отобразить в сравнении с открытой бутылкой, из которой вытекает жидкость через узкое горлышко. Открытое горлышко означает открытую систему, обладающую свойством к непрерывному росту, наращиванию по заданной генами колее. Рост на определённой дистанции ограничен определенными клапанами, то есть при достижении заданных размеров один клапан закрывается и начинается развитие через истекание из другой бутылки, имеющей качественные отличия от предыдущей. Это уже этап не количественного роста, а качественных изменений. Онтогенез — это этапы, или переливание из одной бутылки в другую. Всего бутылок (этапов) = 8. Каждый этап находится в динамике до заданных ограничительных параметров.
Весь геногенез является фиксацией, отображением на клеточном уровне внешнего статуса, состояния, то есть онтофизиса.
Над каждым «Комплексом геногенеза» имеется своя «метка» эпигенеза, который одной «кнопкой» удерживает работу всего комплекса программ. Комплекс — это множество сопряженных программ связанных триггерно, когда одни программы увлекают за собой включение и других. Онтогенез – это комплекс меток эпигенеза, то есть «дорожных столбов». Каждый «столб-метка» определяет особый фенотип онтогенеза. Пока не будет пройден путь реализации по первому столбу до конца, невозможен запуск, проход на втором шлагбауме. Одна эпипрограмма закрывает дорогу к другой. Эпигенез – это смещение с одного статуса стабильного эпигенома на другой, когда становится возможным вскрытие новых гено-программы, что аналогично передвижению ползунка на реостате. Переход ползунка с одного витка на следующий виток возможен только при условии проработки и реализации всего диапазона программ предшествующего витка. За реализацию диапазона отвечают гомеостазы. Переход с одного гомеостаза на другой реализуется с помощью механизмов гомеорезов. Передвижение ползунка обеспечивают «клеточные драйверы».
Онтофизис наиболее существенно проявляет себя на фазе остановки роста организма, а также в условиях перестройки гормонального фона. Любой гормональный сдвиг проявляется в виде метки на эпигеноме. Гормональная зрелость определяет онтофизис. Теоретически онтофизис через эпигенез способствует запуску сенесцента. Через сенесцент клеток онтофизис решает часть своих проблем сохранения стабильности в плане гормезиса. Вне онтофизиса баланс метилирования не нарушается?
Степень проработки метилирования хромосомы определяет как степень её электростатического потенциала, так и степень её магнитной напряженности и закрутки, от которых в свою очередь зависит работа фермента теломераза и репарирование теломеры.
Сброс времени на биологических часах или таймере, а также снятие онтофизисного эпигенеза, зрелости.
Однозначно это происходит при зачатии и образовании зиготы, то есть слиянии двух гамет. До образования гамет в половых клетках, как и во всех других, тоже происходит накопление времени, сенесцент. Например, гаметы-сперматозоиды происходят из сперматогониальных стволовых клеток, которые, подвергаются самообновлению и дифференцировке с последующим двухцикловыми делениями с образованием гаплоидных сперматид, и фазу спермиогенеза. Только в зиготе ползунок реостата или онтофизисный эпигенез обнуляются. Следовательно, в этом механизме образования зиготы скрыт механизм сбрасывания эпигенеза. Очевидно для «сброса» всех онтофизисных накоплений в ходе онтогенеза имеются некие неизвестные нам еще механизмы. Похоже «секрет» заложен в самом процессе мейоза, когда из диплоидной клетки (2n) образуется четыре дочерние гаплоидные клетки (n), отличные друг от друга и материнской клетки. Только в мейозе возможно осуществить полную очистку от «меток» эпигенеза и вернуть тотипотентность. Возможно, попутно они проходят и через гены факторов Яманаки, то есть это сбрасывание происходит ступенчато, где последняя ступенька находится уже в самой зиготе, где и происходит окончательное сбрасывание. Удивительно как чётко происходит не только обнуление эпигенеза, но и полный возврат в исходное состояние метилирования ДНК, заглушка всех меток на гистоновой рубашке, восстановление электростатики на хроматине, а также работы фермента теломеразы и возвращение теломеры до исходных размеров. Шедеврально! Осталось только разобраться как это осуществляется?
Метионин влияет на экспрессию генов плюрипотентности и уровни белка, связанные с метаболизмом метионина, во взрослых, фетальных и раковых стволовых клетках.
Такой ракурс означает, что метионин может содействовать активированию или восстановлению стволовых клеток, функционирующих на уровней тканей. Но действует он не на все типы стволовых клеток, а только на низкопотентные типа прогениторных, что не обеспечивает полную реювенилизацию клеток, сохраняя линии стареющих клеток. Например в культуре ткани метионин и др. аминокислоты ослабляли активацию звёздчатых клеток печени. Они вызывают остановку клеточного цикла в фазе G1 посредством индукции p21. На более высокий уровень потентности стимулирования нужны уже не нутриенты, а гормоны. Было определено, что с увеличением концентрации метионина в культуральной среде увеличивалась экспрессия ряда генов. Таким образом, метионин играет решающую роль в метаболизме и эпигенетической регуляции в различных группах стволовых клеток.
Необъяснённым является то, что с одной стороны высокие дозы метионина активируют стволовые клетки до уровня плюрипотентности, а с другой стороны они же провоцируют старение. Автор считает, что здесь должен быть еще какой-то триггерный промежуточный фактор, от присутствия которого будет зависеть в каком направлении пойдёт сдвиг. Похоже, что в ювенильных тканях метионин будет способствовать их росту и активировать плюрипотентные клетки, на раковых клетках он тоже будет их провоцировать, а на клетках взрослых организмов, прошедших через онтофизисные перестройки, эти высшие программы будут закрытой дверью для плюрипотентности, но откроются двери для усиления сенесцентирования при избытке метионина. Помехой здесь будет онтофизис, который если бы нам удалось убрать, то смогли бы заново запустить программы реювенилизации. У растений этот эффект зрелости ткани или онтофизис удаётся снять гормоном цитокинин, в этом направлении следует искать и возможности его преодоления для омоложения клеток и у человека.
Метионин запускает выработку гомоцистеина – причины возраст-зависимых анизомалий (заболеваний).
В свою очередь метионин является субстратом для конвертирования в гомоцистеин. Это другая кислота, которая тоже является фактором старения. Чем больше метионина, тем больше и гомоцистеина. Однако, это утверждение правомочно только для взрослых, зрелых организмов, тогда как в детских и молодых организмах всё наоборот и он не преобразуется в столь больших количествах в гомоцистеин по сравнению с возрастными людьми. И этот факт показателен, так как является биохимической основой смены принципов клеточного существования.
Эпифиз в роли онтоавтопилота или онтонавигатора
Эпифиз можно сравнить с авиационным автопилотом, координирующем навигацию во времени. Основным устройством этого прибора является гироскоп. Именно он определяет крен, отклонение от заданного курса или норм и параметров. В нашем случае это подразумевает отклонение от параметров гормезиса, а точнее выход за рамки коридора гормезиса. Гироскоп — это прибор видения величины отклонения, а это означает, что и эпифиз несёт ту же функцию — это орган видения, продвижения, эпифизоскопии, но только не в пространстве, а во времени. На этапе эпифизарной молодости* он непрерывно эскалирует – усиливает свою функцию, но он же первый уходит на инволюцию после начала элевации в гипоталамусе. Взаимодействует он с организмом через гипоталамус, а организм с эпифизом - через вегетативную нервную систему (ВНС). Онтогенез идёт на онтоавтопилоте, то есть корректируется сверху. В этой системе эпифиз ведёт себя как транспозит, а гипоталамус – как контрпозит. Все вместе они ведут нашу Жизненную Силу по Коридору Гормезиса. Но в границах оптимума и гормезиса организм находится на онтоэтапах с 1 по 4, а затем происходит выход за рамки гормезиса, система оси эпифиз-гипоталамус начинает давать гигантский люфт, разболтанность в регулировках, а этот люфт открывает двери к многочисленным анизомалиям*. У эпифиза имеется своя система настройки, в число которой входит и ВНС, в частности баланс или дисфункция между симпатической и парасимпатической её отделом. Очевидно, это возможный рычаг для корректировок эпифиза.
Как только «бутылка» гипоталамуса «наполнится», то есть созревает, он начинает подавлять эпифиз; онтонавигаторная = онтодианная* функция эпифиза начинает гаснуть, сохраняя только циркадианную* функцию, при этом гипоталамус переходит на режим автономии. Но в целом онтодиану, её величину определяет эпифиз, а точнее его транспозитную мощь, потенциал, то есть способность противодействовать контрпозитным воздействиям со стороны гипоталамуса. Транспозиты и контрпозиты – это два крыла эпифиз-гипоталамической оси. Транспозитное крыло определяет длительность эпифизарной молодости на онтодианной линии. Кстати, у человека это крыло одно из самых длинных и длится до 12 лет, тогда как у мелких грызунов оно около пол года. Слабый эпифиз, низкая транспозитная мощь его производительности обеспечит короткую онтодиану – и это обозначит для данного вида животного короткожитель ство, высокая мощь – обеспечит длинную онтодиану, то есть определяет потенциал долгожительства.
Принцип перехода от пойкилотопии к гомойотопии – основа смены метаболизма с анаболического на катаболический тип и перехода на закрытую систему существования
Связана эта смена с возрастным переходом с пойкилотопии (принципа непрерывного роста) на гомойотопию (прекращение, активное торможение роста). Количественные изменения при этом переходят в качественные, когда рост заменяется на усиление зрелости, включая со стороны гипоталамуса и изменения гормонального фона, так и со стороны перехода на более высокую степень зрелости клеток. Это особая фаза онтогенеза, которую мы обозначаем как адальтус. Такое торможение роста связано как с изменением гипоталамической регулировки (гипергипоталамическое доминирование), так и закреплено дополнительно коммитативно на уровне клеток с закрытием возможности непрерывной репликации.
В молодости метионин важен в повышенных количествах, так как обеспечивает мощь вегетирования и здоровья, то есть приносит пользу; при зрелости повышает только степень перезрелости и приводит к обратному действию – эпигенетическим проблемам. Сказывается фактор возраста – онтофизис. Он в свою очередь меняет метаболизм клеток, что является основой для повышения степени зрелости клеток в стадии онтогенеза адальтус. Тип клеток тот же, но статус уже другой. То что раньше было хорошо для организма приобретает противоположное свойства.
Чем глубже мы продвигаемся по фазе адальтус, ускоряя её метионином, тем ближе мы приближаемся к фазе климакса и матуры, за которыми уже просматривается и сенилит.
Тотальная резистентность клеток – главная особенность клеточного сенесцента и сенилита.
Хорошо известно, что с возрастом наши клетки и ткани из-за сенесцента становятся нечувствительными по многим параметрам:
- витаминам: витамины D3 (к нему чувствительность с возрастом снижается в 2 раза, то есть в крови он есть в достаточном количестве, но клетки воспринимают как дефицит), B1, B6, B9, B12 …,
- минералам (магний, цинк, кальций…),
- аминокислотам (таурин…),
- клеточным гормонам (оксид азота…)
- и многое другое.
Клетки попросту выходят за пределы по многим константам гомеостазов и функционируют за их пределами, а организм выходит за пределы коридора гормезиса и тоже работает не в оптимуме. Организм воспринимает эту нечувствительность как дефицит, следствием чего является остеопороз, атерослероз, склонности к диабету II, доброкачественным опухолям как аденома простаты, мастопатия, онкология…
В дальнейшем меняется не только метаболизм клеток, но и их чувствительность к регулировочным сигналам, резистентность, а это уже пролог к сенесценту. Онтофизис усиливает резистентность и запускает сенесцент. Сенесцент это одно из проявлений онтофизиса. Даже в культуре клеток, где тоже снят полностью эффект доминирования одних клеток над другими, проявляется онтофизис и нарастает резистентность. Клетки животных не могут находится постоянно в задержанном эмбриогенезе, это для них тоже стресс, не соответствие требованиям их гомеостазов и программ, что и запускает метилирование и эпигенез.
Гомоцистеин повышает риск проявления атеросклероза, болезни Паркинсона и Альцгеймера…
Итак, метионин с одной стороны повышает степень плюрипотентности и реювенилизации клеток, что должно поощряться, но с другой стороны он же провоцирует сенесцент. Причиной этому онтофизис. Открытая вегетирующая система переходит на принцип закрытой, эпигенетически зрелой, а точнее на онтостадийно перезрелую. Открытая вегетирующая система всегда работает на принципах иммортальности клеток (бессмертности), тогда как закрытая система – это самоограниченная система и существует на принципах мортальности клеток, который реализуется через сенесцент. Следовательно, надо уметь противодействовать онтофизису. Последний в свою очередь зависит от гипергипоталамического доминирования. В основе этого лежат элевационные процессы в нём с последующим изменением нейро-гормонального и гуморального фона. Последний сказывается на зрелости клеток и тканей-мишеней в виде вскрытия в них эпигеномных перестроек ведущих к онтофизису. Имеется ввиду, что под онтофизисом подразумевается сохранение своего приобретённого статуса зрелости клеток, включая сохранения их резистентности, даже в условиях если гипоталамус возвратит свой изначальный уровень сигнальной регулировки как в молодости. Для ясности напомню, что в растительном мире клоны побегов, то есть укорененные черенки сохраняют признаки топофиза даже в условиях когда с них снят фактор апикального доминирования. В этом случае для реювенилизации нужны другие условия, которые может обеспечить гормон цитокинин, вырабатываемый уже в других апексах – корневых. То есть нужен гормон молодости и дедифференциации. У растений долгожителей гормоны зрелости, дифференциации и сенилита уравновешены с гормонами противоположного действия – реювенилизации и дедифференциации, что в комплексе позволяет обеспечить закладку новых точек роста и тем самым неограниченное обновление растения.
В организмах животных тоже должен работать этот же принцип для поддержания непрерывности точек самообновления, но с дополнительным условием механизма замены, с удалением старого на новое. Это особенность ткани животного. Кроме всего, для сохранения статуса ткани нужно создать механизмы состаривания части клеток, а затем еще вывести старые клетки с их сенесцентным секретомом, влияющим негативно на весь организм.
Реализация их была обеспечена с появлением новых механизмов как апоптоз, аутофагия, сенесцент. Через реализацию мортальности этих клеток и их аутолиз можно обеспечить освобождение плацдарма для молодых линий клеток. Этого нет у растений. Таким образом, те же принципы самосохранения у животных работают на новых механизмах, то есть всё регулируется по другому: аналогом матрикального апекса является гипоталамус, а эффекта корневого = радиксального универсального действия на все ткани здесь нет, но имеется множество регуляторных веществ не универсальных, а присущих для каждой отдельной ткани. Вырабатываются они не на уровне гипоталамуса, а в каждом типе ткани свой регулятор, который и активирует только свои точки самовосстановления в каждой ткани. Итак, процитокининподобного радиксального (корневого) гормона здесь нет, но в каждом типе ткани имеются свои цитокины, которые вырабатываются стволовыми клетками ткани. Они по сути являются аналогами радиксальных апексов второго, третьего порядков и разбросаны они по тканям всего организма. В свою очередь гормоны проауксиноподобного типа действия, которые вырабатываются в главных осевых или матрикальных апексах у растений, в организмах животных тоже являются централизованы и вырабатываются в гипоталамусе. Это означает, что стволовые клетки тканей играют роль корней в плане выработки гормонов, а главной осью регулировок становится гипоталамус.
Возрастной крен превращения метионина в гомоцистеин очевидно сопряжен с измененным метаболизмом, который определяет онтофизис и опосредуется через усиление резистентности клеток, включая сниженную чувствительность к витаминам группы В, включая B1, B6, B9, B12 в клетках. Именно они или их дефицит определяет перевод метионина в гомоцистеин. Эти же витамины способны «превращать» гомоцистеин обратно в метионин. Это означает, что с возрастом следует принимать эту группу в завышенных супрафизиологических дозах.
Напомню, что продолжительность жизни деревьев долгожителей, например сосны остистой, объясняется мощью воспроизводства цитокининов их корнями. Это так называемый радиксальный эффект, который позволяет растению преодолеть все возрастные проблемы, болезни, травмы, засухи, то есть дать дереву неисчерпаемую Жизненную Силу. Это означает, что Сила дерева находится не в точках роста или верхних апексах, а в корнях! Тем не менее, следует учитывать, что без длительной мощи работы верхних апексов нижние апексы радиксов не смогут развить свою достаточную мощь. Они взаимоувязаны. Но, тем не менее, основную мощь Жизненной Силе дают нижние радиксальные апексы. Например, у многих деревьев если спилить крону она может быстро восстановиться за счет отрастания новых побегов из спящих почек в основании ствола. Из этого следует, что в организме человека Жизненная Сила зависит в первую очередь не от работы гипоталамуса, а его «радиксальной» составляющей, которая имеется в стволовых клетках всех тканей. Они то и выделяют свой секретом с противовоспалительными и регенераторными цитокинами.
Тем не менее, на примере деревьев уточню, что не у всех у них имеется способность к регенерации ствола после его спила. Например, дерево секвойя вечнозеленая способно легко восстановить новый ствол и крону, тогда как его ближайший родственник и такой же долгожитель секвойядендрон гигантский после спила кроны не способен на такое самовосстановление. Известно, что корневая система у него не менее мощная чем у секвойи, но, тем не менее, ему это не дано. Такая особенность связана с лёгкой способностью закладывать спящие почки в основании ствола. Но уточню, что сосна остистая, у которой известны особи доживающие до 4800 лет, тоже не даёт образование спящих почек, а регенерация у неё происходит из адвентивных и брахиальных почек, то есть по потентности более низкого порядка. Тем не менее, этого вполне достаточно, чтобы обеспечить дереву возможность неограниченного существования. Так что мощь возобновляемости деревьев долгожителей, которые восстанавливают себя независимо от того происходит это из спящих почек или пазушных, в любом случае зависит от мощи стимулирующего радиксального фактора. Он для них основной.
В тоже время присмотримся к рисункам [18 и 19], где изображена секвойя произрастающая в разных условиях. Так на фото, где деревья находятся в условиях слабых почв, имеют все признаки старения в возрасте 60 лет, тогда как деревья растущие в условиях глубоких почв поймы реки в возрасте 500-1000 лет имеют здоровый молодой вид и хороший прирост верхушечных побегов. Следовательно важным условием пролонгирования молодости является сохранение, поддержка мощи радиксов, а у человека его стволовых клеток.
В свете рассмотренных фактов, следует утверждать что и в человеческом организме определяющими его продолжительность жизни будут способности к репарации и регенерации его радиксально-подобных стволовых клеток. К сожалению, его стволовые клетки не обладают таким высоким потенциалом секретома, чтобы постоянно возрождать, преодолевать все невзгоды болезней и анизомалий и обеспечить полную реювенилизацию тканей. Радиксальная составляющая определяет особенности продолжительности его жизни. Человек относится к категории средней продолжительности жизни, но не долгожителям, как некоторые виды черепах и так далее. Чтобы перевести человека в категорию долгожителей необходимо содействовать его радиксальной системе стволовых клеток, путём их стимуляции или усиления путём введения их секретома извне. Это мнение подтверждает и феномен так называемого парабиоза, когда сшивали кровеносную систему молодой и старой мыши, что приводило к частичной неустойчивой реювенилизации старой, но более устойчивого и выраженного сенилитирования молодой. Значит фактор что старости сильнее фактора юности.
Возможен ли выход из состояния старения клеток и организма (сенесцента и сенилита) и причем здесь «метиониновый тупик»?
Анализ показывает, что метионин стимулирует молодые клетки и усиливает их Жизненную Силу и способен продлевать молодость. Но всё зависит от возраста. Предполагаю, что на фазе детства дефицит метионина ускорит старение организма, а его избыточность продлит акселерацию и юность.
В молодости и детстве условия обеспечения всеми необходимыми нутриентами, в том числе и метионином, имеют аналогию с ростом у деревьев. Посмотрите на рис. 18 и 19, где четко видно, что молодые деревья секвойи в возрасте 60 лет приобрели все признаки старения в условиях плохого почвенного питания, тогда как 500-летние на хороших почвах имеют здоровый вид. Явное преждевременное старение, которое по внешним признакам подобно старению 4800 летней сосны остистой на рис. 10. В зрелом возрасте у человека и гомойотопных* животных фактор достаточности или избытка пищевых субстратов приведёт к противоположному эффекту – тоже к старению за счёт подключения противоположно действующих механизмов – mTOR. Это означает, что в ущерб ограничению роста, у животных вынужден запуск противоположных механизмов – старения даже в благоприятных условиях. Это означает, что старение у человека вписано в онтогенез на клеточном уровне.
Итак, на более продвинутых фазах – обратный эффект, то есть избыток ускоряет старение, а дефицит - тормозит. Но скорее всего, метионин в зрелом возрасте не является причиной, механизмом старения, а всего лишь потоком, который ускоряет самостоятельное «движение мельницы старения». В основе этой «мельницы старения» лежит повышение клеточной нечувствительности, митохондриальная дисфункция, эпигенез… Стимулирование нутриентами лишь усилит диспропорции между потребностями и возможностями… Нужно усиливать радикальные возможности клеток, что возможно не с помощью нутриентов, а клеточными гормонами-реювенилизаторами аналогичным цитокининам. Следовательно, нутриентным или субстратным фактором у животных эту систему не раскачать. Метионин сугубо нутриентный фактор, который в молодости может активировать, содействовать основному потоку и стволовым клеткам, но не в зрелости.
Обратим внимание на то, что обратными свойствами обладает слабый дефицит нутриентов, который тормозит старение. То есть, по сути, это тоже рычаг влияния на онтогенез. Но возможности этого рычага тоже ограничены. Очевидно, возможности этого рычага тоже можно расширять. Одним из приёмов может быть применение его в виде циклических мощных дефицитов на фоне стимуляторов стволовых клеток, наподобие цитокининам. То есть, периодически нужно более глубоко усилить механизмы аутофагии и катаболические рычаги метаболизма, а на этом фоне активно применять тканевые цитокины животных.
Но, как известно, в условиях стареющих организмов недостаточно работают стволовые клетки, что и является причиной как сенилита, так и сенесцента. В этих условиях особо следовало бы раскачать стволовые клетки, активировать их для получения эффекта реювенилизации ткани за счет активирования точек самовозобновления. Здесь бы и воспользоваться возможностями метионина по повышению плюрипотентности, но дверь для него закрыта… Очевидно, здесь нужны новые принципы их стимулирования, то есть активирования с помощью метионина, но на фоне мощных сенолитиков ведущих к апоптозу с последующей аутофагической разгрузкой, расчисткой. Помехой являются эпигенез и онтофизис с исходом в сенесцент. В то же время, одновременное сочетание приёмов сенолитиков и реювенилизоторов может противодействовать друг другу. Снять это противоречие, очевидно, поможет метод инфракрасных ванн, который содействует ускорению исхода сенесцентных клеток путём апоптоза. Подробности смотрите в соответствующей главе.
Выход за пределы белкового гомеостаза – один из многих звеньев механизма онтофизиса через сенесцент
Белковый гомеостаз в клетке зависит также и от их правильной пространственной конфигурации и конформации, и поддерживается за счет двух противоположных процессов: механизмов корректной сборки белков (и их последующей стабилизации) и механизмов деградации белков с нарушенной структурой (протеолиз). Очевидно, в качестве компенсационных здесь должны подключаться и механизмы аутофагии. В клетках неизбежно с возрастом запускаются возрастные многоплановые онтофизисные перестройки, среди которых происходит и агрегирование белков, а это одна из причин сенесцента и дегенеративных процессов. В этом гомеостазе участвует система БТШ, которые восстанавливают нарушенную структуру белков, и при стрессе (тепловом или химическом) уровень БТШ в клетке возрастает. Однако синтез БТШ в сенесцентных клетках снижается с возрастом. За инициирование синтеза БТШ в клетках млекопитающих отвечает белок SIRT1, который является ферментом расположенным преимущественно в клеточном ядре и отвечает за деацетилирование факторов транскрипции, влияющие на клеточную регуляцию: реакцию на стрессоры, долголетие. Стареющие клетки имеют меньше SIRT1. Особенность этих сенесцентных клеток в том, что они выделяют особый секретом – химические сигналы определяющие секреторный фенотип (SASP) и определяющих статус сенесцента. Очевидно, особенность клеток млекопитающих в том, что эти клеточные перестройки определяются и закрепляются не просто изменением гормонального фона, что ведёт к запуску сопряженных генопрограмм, как это происходит у растений, но еще и закрепляется коммитативно (с обязательством, внутренним «предписанием»), что и становится причиной лимита Хейфлика и появления феномена сенесцента клеток. Но это коммитативное закрепление происходит на другом уровне – эпигенеза.
Снижение SIRT1 происходит за счёт деградации белка, а не изменений в экспрессии генов. Если это так, то это очень важно. Это означает, что причина деградации белков больше связана с эпигенетическими вторичными перестройками и не является программируемым процессом. А раз это не программа, то нужны более общие и простые методы деэпигенизации*! Подлинная причина снижения SIRT1 не известна. Но автор считает, что этот процесс сопряжен с возрастной онтогенетической перестройкой, которая спускается с верхних уровней регулировок (гипоталамус и др.) и запускает клеточные сенесцентные перестройки.
За какие механизмы активного долголетия отвечает клеточный уровень?
Изменение активности сиртуинов в клетках человека является одним из характерных маркеров старения. Также ряд данных указывает на способность сиртуинов влиять на скорость старения клеток.
Белки-сиртуины являются одними из ключевых регуляторов клеточного метаболизма, функции митохондрий и ремоделирования хроматина.
Старение резидентных стволовых клеток играет значительную роль в возраст-зависимом нарушении функций органов и тканей организма человека.
Ряд данных указывают на их связь с динамикой активации регуляторных элементов генома, таких как энхансеры, ассоциированные с развитием генетической программы старения, и перестройки архитектуры ядра, взаимосвязь с динамикой архитектуры хроматина, возраст-зависимым изменением паттерна метилирования, а также экспрессией генов.
Гормезиорез и гомеорез – причина сенесцента, сенилита через эпигенез и деградацию белков.
Причём верхние регуляторные перестройки ведут к гормезиорезу* и выходу за пределы оптимального коридора гормезиса на поздних этапах онтогенеза, что в свою очередь запускает механизмы клеточных гомеорезов* и выхода их за пределы гомеостазов, то есть работа не на оптимальных режимах, выхода за пределы множества нормоконстант*. Гормезиорез ведёт к гомеорезу – это когда ткани и клетки из-за нейро-гормонального дисбаланса (стадия матуры*, перезрелости, климакса, гипергипоталамического доминирования) попадают в новые условия клеточного существования. При этом эта смена условий для клеток не обеспечивает работу в оптимальном режиме, как следствие нарушается, перенапрягается, а затем ослабевает энергетика, наступает митохондриальная дисфункция, окислительный и нутритивный дисбалансы, и многое другое. Это выводит жизнедеятельность клетки за пределы гомеостаза, она вынуждена запускать мощные компенсационные эпигенетические перестройки. Это позволяет пусть менее качественно, но продлить жизнедеятельность клеток, но в ущерб частичной деградации белков, искажается хроматиновая натяжка и электростатический статус, а с ними и экспрессия генов, которые в клеточных поколениях только наращиваются, а не нивелируются. Клетки заходят в затяжной режим стресса, клеточной рецессии*, когда они становятся резистентными ко многим сигналингам и пищевым субстратам, а для их функционального восстановления нужны уже супрафизиологические, запредельные их дозы, то есть настройка «константных точек реагирования» регулирования находится уже не в оптимуме, а за пределами гомеостаза. Если проще, то это функционирование клеток за пределами гомеостаза выводит клетки из строя и к аресту репликативных программ – итоговый, финальный результат. Сенесцент клеток – это эпигенетическое следствие хронического их нахождения в особых условиях матуры или стресса и провоспаления. В растительном мире такого выхода клеток в репликативный ступор нет, как нет феномена сенесцента, так нет и другой крайности - клеточного эндогенного ракового перерождения. Не оптимальные условия для жизни клеток растений, выходящие за критические пределы, приводят к их усыханию, простому самоотключению из системы организма. Листья деревьев при плохом их обеспечении просто желтеют и отмирают – это их механизм экзогенного старения, внутренней коммитированности у них нет; тогда как организмы животных переходят на длительное сенилитирование, а их ткани и клетки на эндогенный сенесцент, что хорошо видно на рис. 12, где на лице у человека проявлены все симптомы старости, тянущейся многие годы. Только в организмах животных включаются коммитативные* (фиксируемые в потомствах) процессы компенсации в виде эпигенетических перестроек, которые позволяют растягивать существование потомств из дочерних клеток за пределами гомеостаза. Причём, теломерные укорочения хромосом не являются ключом проблемы, а являются следствием блокирования теломеразы в ходе коммитирующего эпигенеза. Сенилит имеет свой резко выраженный фенотип, смотрите рис. 12, в отличие от юности, рис. 13. Сенилит обуславливает сенесцент. Клеточный сенесцент тоже имеет эпигенетическую природу, когда эпигенез глушит гены молодости, что имеет определенную аналогию с химерными устойчивыми аберрациями в растительном мире, когда растения могут приобретать устойчивый новый тип морфогенеза. Например, на рис. 25 показана взрослая ель с фенотипом ювенильности эпигенетической природы. В норме этот фенотип меняется на второй год существования растения, а затем оно приобретает новый фенотип зрелости. Такая аберрация связана не с генетическими мутациями, а с некими эпигенетическими причинами. Из рис. 25 видно, что при изменении гормонального фона в апикальной почке (возможно связано с гормоном роста гиббереллином) произошла смена фенотипа с ювенильного на зрелый. Очевидно, и в организмах животных сенесцент клеток является такой же эпигенетической аберрацией, но с переходом не на фенотип ювенильности, а на фенотип сенилита. Причем, особенностью животных является то, что при изменении гормонального фона со старого на молодой (смотрите раздел по парабиозу мышей, когда сшивают кровоток старой и молодой особи) обратные изменения на уровне сенесцента клеток почти не происходят, что связано с коммитативным эпигенезом.

Рис. 25. Внешний вид хвойного дерева ели, сохранившего во взрослом состоянии свой ювенильный фенотип, когда побеги на нём имеют признаки присущие только 1-летним сеянцам. Растение является клоновым спуром с эпигенетической аномалией, которая и позволяет ему сохранять ювенильность у всего клонового потомства. На вершине одного из таких растений «выбились» нормальные побеги, то есть произошло возобладание видового фенотипа за счёт активирования программ взрослости в меристемах почек, при этом эпигенетическая «мутация», сохранявшая морфогенез ювенильности, перестала быть доминирующей. Произошла смена гормонального фона, а с ним и возврат фенотипа.

Рис. 26. На фото представлено лишь одно растение Вероники Андерсона, но с резко выраженными различиями на разных побегах (диморфизм в пределах одной особи), что создаёт впечатление, что это разные растения. Причём побеги с бело-окаймлёнными листьями являются спуром, который размножают как вегетативный клон с декоративными особенностями; но на заднем плане видны «выщепившиеся» побеги с обычными зелёными листьями, то есть вернувшиеся в нормальное исходное растение, в котором образовался спур. В природе очень часто находят такие спуры у самых различных кустарниковых культур. Такие спуры не являются в чистом виде геномутацией, а всего лишь эпигенетической аберрацией, которые при изменении в соотношении фитогормонов могут снять своё доминантное влияние и растение возвращает свой исходный внешний вид. При геномутациях нет возврата в исходное состояние и растение становится новым видом, тогда как эпимутации обратимы, возможен возврат в исходное состояние, что связано со сменой гормонального фона. Пример с данным спуром приведён с целью облегчения понимания онтофизисной* стабильности - причины клеточного сенесцента и синдромокомплекса внешних признаков возрастных старческих изменений и анизомалий* (возраст-зависимых болезней) в организме человека, которые тоже имеют эпигенетическую природу и основаны на нарушении метилирования некоторых генов, что объясняет их экспрессорные или блокирующие возможности. Фактором экспрессором для перехода в стадии зрелости, климакса и сенилита является не прямое переключение генопрограмм в ходе онтогенеза, а вторичные навязанные изменения в эпигененоме. К этому их предрасполагает гомойотопный* статус организма, чрезмерное гипергипоталамическое доминирование и слабая возможность клеточных систем в точках самовосстановления для полного самовозобновления.

Рис. 27. Для сравнения приведён рисунок с внешним ювенильным видом 1- и 2-летних сеянцев ели, имеющих полное морфологическое сходство ювенильного фенотипа с неотеническим* клоном.
Клетки становятся менее чувствительными, требовательными, но зато более живучими в аномальных условиях, и при этом создают анизомалийную* предрасположенность, причину для нарушений на уровне гормезиса. Так открывается специфическая для животных способность к сенилиту (качественное и функциональное изменение, перерождение ткани и клеток), создаются условия для возрастзависимых заболеваний животных. Таких качественных перерождений нет в растительном мире, даже у деревьев которым за тысячу лет, прирост побегов ничем внешне не отличается от таковых у молодых растений. Изменения у деревьев только количественные. Присмотритесь к рис. 10, где изображена сосна остистая возрастом 4800 лет и заметите, что прирост побегов совершенно идентичен молодым растениям и на них нет признаков старения. Растения или древесные виды стареют либо целиком на уровне всего организма, либо не стареют вообще. Старение у растений нисходит только с верхних этажей регулировок Такая возможность качественных перерождений открывается только в клетках животного типа. Да, у растений тоже может наступить матура, то есть генеративная перезрелость, когда оно чрезмерно закладывает плоды или шишки, что может привести растение к истощению и даже к его гибели, но это всё равно регулируется на верхнем гормональном уровне. У животных матура ![]() определяет сенесцент, а сенесцент
определяет сенесцент, а сенесцент ![]() определяет сенилит. Поэтому старение в животном мире движется с двух направлений: сверху
определяет сенилит. Поэтому старение в животном мире движется с двух направлений: сверху![]() и
и ![]() снизу. Сенилит
снизу. Сенилит![]() сенесцент.
сенесцент.
Итак, можно выделить два основных уровня проявления единого процесса старения: верхний - на уровне общеорганизменных нейрогормональных регулировок – сенилит, задающий тон и направленность всему организму и зависящий от заложенного потенциала возможностей гипоталамуса; и нижний – сенесцент на уровне клеток и тканей, несущий коммитированные ограничения на уровне эпигенеза. Все они на разных уровнях имеют в своей основе разные механизмы, но в то же время являются разноуровневыми проявлениями Единой Матричной Системы. Эти потоки сливаются в единый – синдромокомплекс геронтоса*, который является анизомалийным полем, единой базой для всего спектра, букета анизомалийных возрастных проявлений или по другому симптомокомплекса болезней старости. Геронтос – это единство сенесцент ![]() сенилит.
сенилит.
Следовательно, и методы противодействия должны быть двусторонними. Но первичен при этом сенилит, а точнее его предшественник – матура. Матура – это последний этап адальтуса*. Адальтус исходит из юности.
Кстати, этот же феномен клеточной рецессии и репликативного ступора может вызвать через эпигенез не только онтофизис, но и многочисленные другие факторы клеточного стресса, в том числе и содержание их вне условий организма культуры ткани. Причин старения клеток много, но механизм реализации один - исход в репликативный сенесцент. Он сопряжён с конформационной (пространственной) деградацией структуры у ряда белков, нечто похожее на денатурацию белков при их нагревании. Белки становятся нерабочими. В норме этому должны противостоять SIRT1 (отвечают за выработку БТШ, регулируют процессы старения, транскрипции, апоптоза и сопротивляемость стрессу и голоданию, отвечают за продолжительность жизни некоторых организмов деацетилирующий фактор транскрипции), но на пути этому становится эпигенез. Он меняет хроматиновые структуры, в том числе блокирует теломеразу. Очевидно, эпигенез можно разделить на приобретённый, то есть связанный с внешними факторами, а также эпигенез, к которому есть онтогенетическая предрасположенность, он становится неизбежным, маскируется под программативный процесс. Процесс развивается на платформе эпигенеза, но причиной ему являются морфогенетические онтогенетические перестройки.
Неотения – путь к разгадке реювенилизации.
К сожалению, этап юности всего лишь промежуточный и обязательно скатывается в адальтус. Возможно, это связано с тем, что природа не может долго удерживать этот статус, так как он связан с непрерывным ростом, что противоречит принципам гомойотопии*. Но все же этот принцип гомойотопии можно поддерживать долго и на фазе юности, но для этого нужны особые механизмы поддерживания неотении*. Примером сохранения такой длительной неотении являются животные голые землекопы, которые сохраняют многие признаки девственности даже при большом возрасте. Многие самцы из одного семейства слабо половозрелы, это не состоявшиеся еще «юноши», но становятся таковыми если исчезают половозрелые. В принципе известны случаи их доживания до 27 лет, когда аналогичные по размерам животные как мыши живут 2-3 года. Неотения здесь определена на верхнем уровне регулировок, а именно на гипоталамическом. Гипоталамус у них недозревший, не продвинутый элевационными механизмами и равноценен таковым у особей детско-юношеского возраста. Это означает, что нет сильного опускания верхнего процесса на клеточный уровень с подключением сенесцента. В условиях сохранения ювенильности организма сенесцент клеток исключается. Всё замедленно, приторможено. Длительное состояние ювенильности у них не допускает матуру и сенилит. Даже отсутствие волосяного покрова у них в зрелом возрасте напоминает отсутствие пубертата, то есть оволоснение у человека. Очевидно эти признаки отсутствия пубертата на коже имеются и в гипоталамусе, где тоже длительно сохраняется отсутствие пубертатных гормональных настроек. Причем состояние допубертатного статуса, то есть отсутствие оволоснения у них закреплено генетически, так как даже у половозрелых особей шерсть отсутствует.
Инфракрасные ванны - способ преодоления клеточного сенесцента через механизмы Белков Теплового Шока (БТШ)
Инфракрасное излучение - способ активизации белков молодости - сиртуинов, включая SIRT1, противодействует деградации белков и сенесценту через усиление выработки БТШ.
БТШ являются универсальными молекулярными шаперонами или упрощенно «расчёсками» восстанавливающими конформацию белков. Автор выдвинул концепцию, что клеточное старение сцеплено с онтофизисом и является клеточной коммитацией (предписанием), которое затем реализуется через те же механизмы, что и клеточный стресс. Но в отличие от стресса, когда внешние факторы можно устранить и тем самым снять его последствия, при коммитации эти же процессы запускаются изнутри эпигенетически. По сути онтофизис – это наведённый и закреплённый в клетках процесс как результат онтогенеза.
Любой стресс в клетке (тепловой, химический или изоляция от сообщества, или нехватка нутриентов) приводит к нарушениям в структуре белков. В норме такие «поломанные» белки должны:
- либо утилизироваться в протеасомах/аутофагосомах, чему можно содействовать с помощью методов аутофагии при интервальном голодании.
Если в системах утилизации-сборки происходит сбой, то белки агрегируются и ухудшают работу клетки. Это способствует старению и развитию дегенерации клеток.
Польза инфракрасного тепла заключается в активизации и оздоровлении всех клеток организма.
Исследование показало значительное улучшение внешнего вида и эластичности кожи после 12 недель сауны. У участников уменьшились морщины и "гусиные лапки", а также улучшился тонус, чистота, упругость и мягкость кожи.
Инфракрасные лучи повышают уровень эндорфинов, что способствует позитивному настроению и безмятежному расслаблению, стимулируют выработку BDNF (нейротрофического фактора головного мозга), который защищает новые нейроны в головном мозге от повреждения из-за стресса.
Проникновение ближнего инфракрасного излучения глубоко в ткани организма может в четыре раза улучшить здоровье клеток и рост тканей. Такое излучение стимулирует выработку белых кровяных телец, рост коллагена и регенерацию клеток за счет увеличения клеточной энергии.
Нарушение белкового гомеостаза и усиление провоспалительных процессов провоцируют конечные продукты гликирования (неферментативной реакции присоединения углеводов к аминокислотам в составе белка).
Высокий уровень конечных продуктов гликирования (advanced glycolation products, AGEs) в тканях вызывает окислительный стресс и воспалительные процессы, так как AGEs связываются с поверхностными клеточными рецепторами, запускают воспалительный NF-kB-сигнальный путь, а также изменяют структуру и функции белков. Снижение количества AGEs в пище замедляет развитие хронических болезней и старения у животных и, видимо, у человека. Овощи, фрукты, зерновые, бобовые, молоко и хлеб содержат мало AGEs, в то время как в твердых сырах, говядине, свинине и птице количество AGEs высоко.
Красный и ближний инфракрасный свет обеспечивают максимальное проникновение в клетки.
Здесь он используется для увеличения выработки клеточной энергии. Цитохром с-оксидаза, фермент действует как фотоакцептор красного света. Как только ферменты цитохрома в митохондриях всех ваших клеток поглощают красный и ближний инфракрасный свет, выработка энергии усиливается, митохондрии улучшают работу.
С увеличением энергии каждая клетка, орган и ткань в организме работают эффективнее.
Терапия красным и ближним инфракрасным светом стимулирует высвобождение стволовых клеток. Они мигрируют в место повреждения тканей и помогают восстанавливать и регенерировать кожу.
Красный и ближний инфракрасный свет уменьшают воспаление кожи.
Воздействие на кожу красного и ближнего инфракрасного света улучшает кровоток по всему телу, что уменьшает острое и хроническое воспаление и ускоряет заживление. Одним из наиболее воспроизводимых эффектов фотобиомодуляции является общее уменьшение воспаления.
Также красный свет для кожи проявляет мощное антиоксидантное действие, которое защищает её и весь организм в целом от многих экологических угроз, с которыми мы сталкиваемся ежедневно, включая яды в продуктах питания, средствах личной гигиены и ионизирующее излучение.
Противовоспалительные, антиоксидантные и улучшающие кровоток свойства красного света являются одними из ключевых свойств, влияющих на оздоровительный, омолаживающий эффект для кожи.
Коллаген — это белковая матрица, которая обеспечивает прочность кожи и буквально всего тела. Он содержится в шаровидных суставах тела, которые также получают пользу от красного света. Получать коллаген можно не только из пищевых источников, но также путём использования красного света, чтобы стимулировать организм к его естественному производству.
Лечения естественным светом, включая определенные длины волн ближнего инфракрасного света позволило увеличить синтез коллагена, выработка которого с возрастом существенно снижается.
Доказано, что красный свет успешно можно использовать для стимуляции более быстрой регенерации кожи и увеличения выработки естественного коллагена.
Показано, что шапероны работают через эпигенез, а значит могут противодействовать сенесценту, способствовать иммунитету и долголетию.
Регулярное использование инфракрасной сауны широкого спектра действия использует лучистое тепло этих световых волн для увеличения производства белков теплового шока, тем самым создавая внутриклеточное сообщество, богатое структурированными белками и свободное от вредных свободных радикалов.
Направленность физиологического действия выработки БТШ связана с предотвращением неврологических дегенеративных заболеваний, улучшением здоровья сердечно-сосудистой системы, а также содействует более длительной периодической ремиссии аутоиммунных заболеваний.
Для чего служат Белки Теплового Шока?
Эти «молекулярные сопровождающие» играют важную роль в сохранении нашего физического, а также эмоционального здоровья и благополучия.
Терапия через БТШ.
Когда термическому стрессу подвергаются организмы от плодовых мушек до человека, происходит следующее: все эти наблюдаемые органические структуры начинают производить Белки Теплового Шока. Когда БТШ создаются в ответ на температурный стресс во внешней среде, они оказывают решающее воздействие на межклеточную среду во всех типах организмов.
БТШ дают возможность каждому организму «активировать» экспрессию генов. Способность организма вырабатывать БТШ тем самым инициирует адаптивную способность межклеточных сообществ находить гомеостаз внутри и, в конечном итоге, побеждать неврологические дегенеративные заболевания, такие как болезнь Альцгеймера, БАС и болезнь Паркинсона.
Таким образом, доставка экзогенных БТШ или стимулирование секреции БТШ окружающими клетками позволило разработать терапевтический подход к лечению нейродегенеративных заболеваний, и признать БТШ в качестве внутриклеточного терапевтического средства, модулятора нейродегенеративных заболеваний.
Что такое Белки Теплового Шока?
Все они классифицируются по их молекулярной массе и их конкретным внутриклеточным функциям, их очень много. При этом их всех называют «молекулярными шаперонами», так как они служат для повторной стабилизации, реорганизации и омоложения межклеточного порядка. Рассматрение БТШ в качестве шаперонов оправдано, так как они служат для сопровождения внутриклеточной транслокации белков.
БТШ играют важную роль в поддержании эффективности любой системы или систем организмов. Показано, что увеличение количества БТШ в физиологии человека способствует предотвращению серьезных неврологических состояний, а также других хронических аутоиммунных заболеваний.
БТШ вырабатываются, когда организмы подвергаются колебаниям температуры, достаточно экстремальным, чтобы выйти за пределы привычной температуры.
БТШ вырабатываются не только в инфракрасной сауне, но и при холодовых процедурах.
Польза для здоровья от увеличения количества БТШ в организме:
- повышенный иммунный ответ,
- сокращение свободных радикалов,
- более быстрое восстановление мышц,
- уменьшение выраженности биологического возраста,
- профилактика неврологических и дегенеративных заболеваний.
Инфракрасные ванны стимулируют выработку БТШ внутри клеток. Здоровье и благополучие физиологии человека зависят от поддержания трехмерного качества белков. Когда белки теряют целостность своей формы или бесцельно задерживаются между клетками, негативные последствия для здоровья становятся экстремальными.
БТШ играют очень важную роль в физиологии человека в отношении их функции молекулярных шаперонов: для обеспечения того, чтобы белки и пептиды сохраняли свою форму и назначение.
Все показатели здоровья и хорошего самочувствия улучшаются, когда количество производства БТШ увеличивается.
В принципе многие тепловые процедуры способствуют выработке БТШ, но тем не менее, сауны с инфракрасным излучением имеют ряд преимуществ для здоровья. Они проявляют более выраженный клеточный ответ на инфракрасные волны, усиливает оксигенацию кровотока.
Считают что работают эти волны через митохондрии, которые становятся более активными. При этом происходит как усиление выработки оксида азота митохондриями, так и улучшение секрецией БТШ. Всё это сопровождается улучшением самочувствия.
В целом молекулярные шапероны и БТШ окажут влияние на увеличение продолжительности и качества жизни.
Оптимальное время суток для пользования сауной?
Посещение сауны почти всегда полезно для улучшения общего состояния здоровья. Некоторые данные указывают на целесообразность посещения сауны утрам, что способствует улучшению умственной концентрации в течение дня.
Регулярное посещение сауны оказывает положительное противодействие старению мозга путём активации Nrf2.
Комплекс условий нужный для адениновых индукторов, чтобы они смогли сработать в направлении реювенилизации
Адениновые индукторы могут пойти по разным путям потентного клеточного пробуждения, инициирования. Существует не менее 5 этажей потентности, где самая высокая открывается на уровне тотипотентности, а самая низкая – на уровне прогениторных клеток. Более качественная и глубокая реювенилизация происходит при возврате на изначально первичные этажи потентности. Но для этого нужны не просто наличие гормонов-индукторов, а создание целого комплекса факторов и условий для глубокой реювенилизации ткани. Для этого нужно наличие не одного гормона или индуктора, а целая система гормонального и метаболического фона, специальный системный фон.
В отличие от растений, где на первом плане стоят институированные механизмы регулировок онтогенеза, в организмах животных на первый план выходит изначальное коммитирование клеток, предначертание, закрепление плана и способа развития, предписание степени возможности возврата уровня потентности. Это на много усугубляет, ограничивает возможности гормонального регулирования процессами ювенильности и зрелости и эти функции регулировки у них переданы с верхних этажей на нижние – клеточные. Поэтому здесь важны не сами гормоны молодости, а именно внутриклеточные регуляторы. Очевидно таковыми и являются микроРНК.
Напомню, что яйца кур состоят преимущественно из белков и липидов, но практически отсутствуют углеводы. И это не спроста. Это наводит на мысль, что для инициации реювенилизационных программ нужно ограничить углеводный обмен, ограничить количество калорий, но насытить организм белками и нуклеиновыми кислотами. Этим самым мы ограничим возможности эндогенных внешних этажей регулировок, но усилим клеточные первичные этажи регулировок.
Следовательно на этот период надо снизить калорийность до минимально возможного – 600 - 900 кал/сутки, чтобы не возбудить процессы роста, не усилить гипоталамическое доминирование, но инициировать репаративно-регенеративные процессы, то есть сомато-стем-доминирование, когда представлено больше возможностей саморегулировки именно клеточному уровню.
Особенность состава липидов. Они тоже должны быть не все подряд, а преимущественно те, которые имеются в желтках яиц – фосфолипиды – лецитины, а не насыщенные жиры типа сала.
Особенность состава белков. Белки строятся из аминокислот и их состав должен быть приближен к яичному, то есть тех, которые особенно важны на стадии становления организма. Поэтому предложено использовать живой белок яиц как дополнительный ингредиент в программах реювенилизации. Таким образом, приём живых пробужденных яиц не просто дополняет приём блюда и молок рыб, а является обеспечивающим фоном, на котором может проявиться эффект реювенилизации клеток и тканей. Подробности об яичной диете можно познакомиться в статье Г.А. Гарбузова: «Яйца – овотерапия, лучшая диета, которая сдерживает старение» [6].
Оздоравливающие возможности рыбных молок
Молоки представляют собой семенную жидкость рыб-самцов. Вкус молок лососевых рыб очень специфичен, потому не все люди их охотно употребляют. В большинстве случаев это обусловлено неосведомленностью о пищевых свойствах данного продукта и неумением его правильно приготовить.
Главное пищевое достоинство молок лососевых рыб – это наличие в них жиров с большим содержанием полиненасыщенных кислот. Продукт способствует нормализации работы сосудистой системы, выступает профилактическим средством таких болезней как закупорка сосудов, инсульт и инфаркт. Входящие в его состав компоненты усиливают действие препаратов для диабетиков и улучшают мозговую деятельность. Молоки содержат полноценный набор витаминов, как А, Е, РР и С, а также группа витаминов В, и весь спектр нужных минералов. 100 граммов рыбьих молок закрывают практически полностью суточную потребность человека в витамине В12, который отвечает за обезвреживание гомоцистеина, нормальную работу нервной системы, профилактику образования атеросклероза и тромбов.
Содержание жира и углеводов в молоках незначительно, но зато многообразие аминокислот впечатляет. Молоки являются источником жизненно важных аминокислот. Среди них следует выделить глицин, который часто используют для стимулирования деятельности мозговых клеток, а также для лечения каких-либо неврологических заболеваний. При употреблении молок у человека повышается иммунитет, а также подавляются воспалительные процессы в организме. Более того, такие вещества способствуют быстрым заживлениям ран (язвочек) и улучшению кроветворных функций в организме. Это натуральный продукт, который обеспечивает всеми необходимыми элементами питания из пищи без применения искусственных добавок.
Калорийность 99 ккал на 100 гр.
Блюда из молок.

Рис. 28. Натуральные сырые молоки.

Рис. 29. Блюдо приготовленное из молок на пару.

Рис. 30. Паштет из молок.
Возможное побочное действие рыбных молок
Очевидно следует ожидать, что поскольку эти молоки содержат избыточно нуклеотиды, то значит и аденины, которые являются одновременно и пуринами – источника мочевой кислоты. Она скапливается в хрящах, сосудах, суставах и тканях. Это может стать причиной подагры, поражающей опорно-двигательный аппарат, и почечной недостаточности. Если в организме человека содержание пуринов значительно превышает норму, это препятствует их нормальному выведению и приводит к ухудшению самочувствия.
Для взрослого человека нормой считается 650-1000 миллиграммов пуринов в день. Первым признаком переизбытка вещества является лёгкая аллергическая реакция (высыпания на коже).
Способ преодоления. Поскольку пурины образуют в определенных условиях натриевые соли, которые и обладают способностью окладываться в суставах, то предлагается:
- ограничить приём соли кухонной,
- но увеличить приём альтернативной калиевой соли,
- время от времени устраивать разгрузочные дни – умеренное голодание, а лучше интервальное питание с целью усилить аутофагию и изъятия избыточных отложений,
- одновременно принимать «живые» напитки, которые имеют минусовой заряд и не дают возможности выпадать в осадок солям пуринов.
Какая аналогия в действии цитокининов и гидролизата плаценты?
Плацентарная жидкость – экстракт плаценты – средство от 1000 возрастных болезней и омолаживания.
Применение плацентарных экстрактов кроме целебного действия имеет элементы реювенилизации. Основные компоненты их действия связаны с содержанием набора фетальных белков (белки поздних этапов эмбриогенеза, часть из которых ведут к стимулированию процесса «распаковки» генетических программ развития эмбриона, а другая часть обеспечивает эмбриогенез как на стадии закладки зачатков, так и последующее разворачивание из зачатков тканей эмбриона). Таким образом, они принципиально отличаются от действия тотипотентных белков, которые появляются на стадии зиготы и проявляют универсальную = абсолютную регенерацию. При этом стимулы к действию исходят исключительно изнутри первичных клеток без всяких внешних сигнальных корректировок. При этом клетки такого бластомера способны бесконечное число раз восстанавливать недостающее количество клеток в случае удаления некоторых из них. Таким образом механизм сохранения гомеостаза на этом этапе развития идеален.
Поздний эмбриогенез обеспечивается с помощью работы плюрипотентных и мультипотентных генов, присущих и стволовым клеткам. Задействование этих фетальных белков способно делать среди зрелых и стареющих линий клеток частично то же что и в эмбрионах. Они же способны оживить пулы генов унипотентных клеток = «листьев» = дифференцированных клеток.
Очевидно, что филогенетически механизмы и белки обеспечивающие становление эмбриогенеза едины среди мира животных и имеют идентичность плюрипотентных и мультипотентных химических индукторов. Это означает что эти белки можно получать не только от особей человеческого вида, но и различных других.
Любое лечение предусматривает временное воздействие, тогда как оздоровительные или омолаживающие методики подразумевают практически постоянное применение.
Плацентарный экстракт не обладает таковыми абсолютными потенциями. Но, тем не менее, тоже имеет огромное скрытый спектр других возможностей, способный заводить регенерацию почти во всех видах тканей организма.
В первую очередь это мощный активатор становления, восстановления и реконструкции сосудистого русла. Именно плацента обладает свойствами обеспечивающими растущему плоду становление сосудистой системы. Специфика млекопитающих, у которых плод развивается в матке, обусловлена необходимостью воспроизводства инвазийной ткани плода, обильно заполняемой кровеносными сосудами, которые через пуповину сливаются в единую кровеносную систему в стенке матки, переправляя, переориентируя движение кислорода и питательных вещества из организма матери на питание растущего плода. Следовательно, плод = эмбрион создает плацентарную среду вокруг себя, которая стимулирует гормонально матку. При этом стимулируется не только плод, но и матка, да и весь организм женщины. Улучшаются трофика, поставка кислорода, перестраивается гормональный фон и гормонообеспечение плода.
Естественно ожидать, что эта же жидкость окажет омолаживающее действие на любой организм. С возрастом как известно наша микрокапиллярная система теряет до 50% капилляров и это одна из причин тотального нарушения трофики и старения. Здесь особо важен аналогичный стимул. Автономные системы саморегулировки и поддержанию гомеостазов по восстановлению этого русла с возрастом по ряду причин все больше и больше слабеют, теряют жизненную силу, что связано как с болезнями, так и с запуском механизмов инволюции = сенилита.
Недифференцированные клетки плаценты и плода также попадают в кровоток матери и могут даже превратиться в часть её обычных тканей тела – этот процесс называют микрохимеризмом. Процесс можно расценивать как донорство стволовых клеток и их индукторов.
Особо выражено гидролизат плаценты (ГП) действует на сосудистую систему. Люди которые пользуются им регулярно надолго или вовсе отодвигают эти проблемы.
Очевидно в первую очередь это связано с регенеративным воздействием на эндотелиальный слой сосудов. Именно здесь в эндотелии начинаются все возрастные проблемы в сосудах, в том числе и замедленное восстановление микрокапиллярного русла, с его постоянным «пересыханием», а также нарушением капиллярной трофики крупных сосудов, где первично начинаются пожары воспалительных процессов, приводящие к эндотелиальной сосудистой недостаточности с последующими дегенеративными процессами как атеросклероз и артериосклероз.
Также ГП активно воздействует на восстановление возрастных проблем в суставах, как артриты, остеохондроз, что тоже связано с активацией регенеративной способности любых тканей, ну и конечно с восстановлением нарушенной капиллярной сети. В принципе все возрастные изменения в любых тканях однотипны – это ухудшение кровоснабжения, уменьшения капиллярной сети, проблемы эндотелия в крупных сосудах сопряженными с хроническими возрастными провоспалительными проблемами в них приводящих к атеросклерозу и склерозу, а также возрастного затухания регенеративных и репаративных способностей, что связано с возрастными ограничениями, исчерпания лимита возможностей к митозу клеток. Снижение клеточной активности – это встречный процесс, идущий снизу с уровня клеток и связан с клеточным лимитом Хэйфлика. ГП действует и на этот поток возрастных процессов.
Неудивительно, что ГП окажет своё положительное действие в любой ткани, включая и нервную, что улучшит её функциональную работоспособность, а значит улучшит память, сон, повысит работоспособность, ослабит депрессию, уменьшит уставаемость, повысит зрение, слух, улучшает самочувствие, гармонизирует половую дисфункцию.
Заметное улучшение окажет на качестве и состоянии кожи, вплоть до уменьшения возрастной пигментации кожи, уменьшения полипов, выпадение волос, снижая симптомы псориаза, нейродермитов, герпеса, печени (гепатит, цирроз), иммунитета, уменьшает симптомы гормональных перекосов...
Так можно пройтись по всем 16 функциональным системам организма где ГП окажет своё положительное действие. В целом это увеличит общую продолжительность жизни человека.
Основное действие препарата связано не с витаминами, ферментами и гормонами, а благодаря присутствию только в нём специфических уникальных белков – индукторами, активаторами на уровне генома клеток. По сути их можно назвать стрелочниками перенаправляющими локомотив на путь омоложения, предотвращая возрастное сдерживание и угасание клеток, включая их самообновляемость.
Цитокины в организме животных: что общего с цитокининами растений?
Цитокины и регенерация.
Цитокины – активные участники репаративных процессов в различных тканях. В этих процессах участвуют множество из известных цитокинов (ИЛ-1, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10, ИЛ-18, ТФР, ФРФ, РФЭ, ФРК, РDCF). Они способны оказывать прямое влияние на клетки и ткани или опосредованное через другие молекулы.
Большинству из цитокинов присуща плейотропность, они способны выступать в качестве активирующих и ростовых факторов для клеток, переводить их в состояние, в котором они способны воспринимать митотические импульсы. Известно, что активированные лимфоциты способны повышать в 2-3 раза митотическую активность самых разнообразных клеток (клеток печени, эпителиоцитов, клеток эндотелия сосудов, миоцитов и др.). Установлено, что ИЛ-1 способен стимулировать пролиферацию фибробластов, основных клеточных элементов соединительной ткани и продукцию ими коллагена, пролиферацию эндотелиальных клеток. ИЛ-6, ИЛ-10 усиливают рост эндотелия и новых кровеносных сосудов. ИЛ-18 усиливает репаративные процессы в хряще, повышает активность хондроцитов, стимулирует выработку ими гликозаминогликанов. Трансформирующий фактор роста бетта (ТФР), основной фактор роста фибробластов (ФРФ), ростовой фактор эндотелия (РФЭ) (VEGF) и МСР-1 способствуют ангиогенезу, откладыванию коллагена и белков во внеклеточном матриксе тканей. Фактор роста кератиноцитов (ФРК) усиливает реэпителизацию тканей, дифференцировку фибробластов, что в итоге стимулирует заживление ран. Тромбоцитарный фактор роста (PDGF) обладает митогенной активностью в отношении гладкой мускулатуры. Он стимулирует выработку моноцитами и макрофагами других ростовых факторов (ТФР).
Имеющиеся клинические данные свидетельствуют о том, что заживление ран, репаративные процессы в различных органах и тканях и восстановление их структурной и функциональной полноценности протекает намного активнее у лиц с высокой иммунореактивностью и способностью продуцировать цитокины, чем у лиц с депрессией иммунных реакций.
Цитокины - проводники молодости.
Выделяя и выращивая в лабораторных условиях стволовые клетки, получаемые из экстрактов эмбрионов и плаценты, специалисты действительно вплотную приблизились к истине, но помешало одно существенное обстоятельство. Оказалось, что стволовые клетки, обладая уникальной способностью продуцировать новые источники восстановления в организме, проходя через обычную инъекционную иглу, погибают или, теряя свои свойства, не успевают доносить до проблемной зоны необходимую информацию.
В связи с этим несложно предположить, насколько революционным событием в мире клеточной биологии стало открытие цитокинов - регуляторов клеточной активности и клеточных факторов роста. Эти вещества не только способствуют выделению в кровь большого количества стволовых клеток, но, главное, локализуют их там, где это требуется в данной ситуации. Управляя процессами их размножения, миграции и жизнеспособности, цитокины как добросовестные проводники доносят стволовые клетки до пораженных систем, побуждая их к дальнейшему воспроизведению, а также к синтезу собственного коллагена — основного стройматериала кожи.
Эффективность цитокинов в омоложении организма, а также заживлении кожных ран подтверждена. Когда производство активных в отношении стволовых клеток цитокинов было налажено, их начали широко применять в пластической хирургии для лечения травм, а после получения положительных результатов - и в омолаживающей клеточной терапии.
Сегодня метод биологического омоложения успешно используют как для улучшения состояния кожи, так и с целью общего омоложения организма. После инъекции цитокинов, которые немедленно стимулируют размножение стволовых клеток, пациент обретает совершенно новое качество жизни. Ведь в этом процессе задействованы все системы, в том числе и гормональный фон.
Визуальные признаки молодости проявляются прямо на глазах и с каждым днем становятся все более заметными. Отступает большинство хронических болезней, существенно повышается общий тонус, а также либидо и потенция у мужчин. Еще одно ценное качество цитокинов - рост сосудов на фоне оптимизации сердечных показателей. Кардиологи с помощью этой методики добиваются нормализации кровоснабжения миокарда при ишемической болезни сердца.
Для достижения оптимального результата процедуру биологического омоложения посредством цитокинов разделяют на несколько этапов. Длительность курса может быть разной.
Следующий этап — инъекционное введение цитокинов непосредственно в организм. Чтобы использовать цитокины для активизации размножения фибробластов и синтеза собственного коллагена, их выделяют из крови пациента совместно со стволовыми клетками. Иными словами, организм учится омолаживать себя самостоятельно, но насколько быстро это произойдет, зависит только от индивидуальных особенностей человека.
Возможности мелатонина в реювенилизации
Главный гормон эпифиза мелатонин обладает противовоспалительным действием, снижая концентрацию белка NF-kB, он снижает весь уровень усталости и воспаления, накопившиеся за день в любых тканях. Сенилит и сенесцент – это персистирующие провоспаления на разных этажах регулировок организма. С возрастом после 10 лет уровень мелатонина в эпифизе начинает многократно раз снижаться и в старости его уровень падает 10-ти кратно. Напомню, что гипоталамус имеет особенность на последних этапах онтогенеза - усиливать провоспалительное состояние и тем самым продвигать этап старения = сенилита организма. Долгожительство животного зависит от запаса прочности, ресурсов гипоталамуса. Напомню, что с возрастом происходит нарастание нечувствительности гипоталамуса к ряду сигналов = резистентность. Гипоталамус стареет быстрее всех, поэтому противодействием, буфером гасящим этот пожар, должен быть эпифиз с его мелатонином. Но функция эпифиза тоже угасает и причиной тому является усиление доминантной зрелости гипоталамуса. Очевидно, если бы гипоталамус не переходил некие рамки зрелости, то это не ингибировало бы эпифиз. Очевидно, это и происходит в случае с гипофизарным нанизмом, когда тормозится появление признаков пубертата и адальтуса, но пролонгируется ювенал. Из незрелого органа гипоталамус «наливается», наполняется зрелостью. Половозрелость, гиперзрелость начинают доминировать над ювенильностью. «Переключателем» на зрелость становится сам гипоталамус, без его сигнала низы сами не могут запустить репродуктивную зрелость, а затем и климакс и сенилит. Приоритеты меняются в онтогенезе. Если бы мы научились продвигать доминанту эпифиза, не допуская или снимая процесс его созревания, то тем самым смогли бы снять статус гипердоминанты гипоталамуса.
Мелатонин оказывает адаптогенное, седативное, снотворное действие. Нормализует циркадные ритмы. Увеличивает концентрацию гамма-аминомасляной кислоты (ГАМК) и серотонина в среднем мозге и гипоталамусе, изменяет активность пиридоксалькиназы, участвующей в синтезе ГАМК, дофамина и серотонина. Очевидно мелатонин мог бы сгладить те дисбалансы, крены, которые происходят в гипоталамусе с возрастом. Существуют доказательства в пользу важной роли эпифиза в контроле за продолжительностью жизни получены в экспериментах с пересадкой эпифиза и введениями эпифизарных препаратов. После трансплантации эпифизэктомированным старым мышам железы, взятой от молодых доноров той же линии, подобные обладатели «молодого» эпифиза жили гораздо дольше по сравнению с соответствующим контролем. В то же время хроническое введение пептидного препарата железы эпиталамина и мелатонина увеличивало на 20–30% среднюю продолжительность жизни мышей и крыс. Была предпринята попытка в клинических условиях пожилым людям (старше 60 лет) длительное время (в течение 3–10 лет) проводить инъекции эпиталамина. Отмечены субъективное улучшение самочувствия пациентов, а также объективная нормализация биохимических и некоторых иммунологических показателей.
Есть основания предполагать наличие у мелатонина геропротективных свойств, а также предполагать значимость гормонального дефицита в генезе двух наиболее распространенных форм возрастной патологии – нарушении когнитивной деятельности мозга и развитии опухолевого процесса. Факты подтверждают наличие такой связи. Но решающего значения в полной реювенилизации он не оказал. В целом он имеет второстепенное, но не ключевое значение в точках самовосстановления организма (пробуждение стволовых клеток). Напомню, что по нашей концепции таковыми должны быть вещества с цитокининовоподобным действием, то есть обладающим универсальным омолаживающим действием на все типы тканей.
Уточню, что гипоталамус является первой скрипкой в продвижении онтогенеза, а значит и сенилита. От его состояния и мощи зависит скорость наступления сенилита. От состояния гипоталамуса зависит баланс всех медиаторов мозга, которые обеспечивает трофику вегетативной нервной системы и всех тканей мишеней. Нами этот процесс обозначен как медиация тканей через мозг. От состояния мозга зависит трофика тканей. Поэтому нужно поддерживать правильный тонус гипоталамуса, действующего как настраивающий камертон. Поэтому в Комплексной Программе по Омоложению важно применять как мелатонин, так и группу цитокининоподобных веществ. У растений нет нервной системы, поэтому им не нужна группа, веществ корректирующих баланс медиаторов.
Поэтому мелатонин должен быть дополнением к цитокининоподобным веществам. Но напомню, что гипоталамус и эпифиз дряхлеют из-за угасания в них стволовых клеток, а мелатонин – это всего лишь продукт функциональной деятельности циркадной системы, от которой зависит качественная сторона функционирования гипоталамуса. Простой приём мелатонина не омолодит эту систему, а только компенсирует её недостатки, что улучшит нервную трофику. Добавки мелатонина даже в высоких дозах не способны довести до такой степени омоложения, чтобы вызвать образование новых зачаточных тканей, а затем из них процессы регенерации, также ему не по силам снятие эффекта онтостадийности. Мелатонин не способен запустить тотипотентность и плюрипотентность, тогда как цитокинин это реально осуществляет.
Способы применения мелатонина. Профиль безопасности препаратов мелатонина очень высок. Мелатонин не обладает повреждающим действием даже при длительном применении в огромных дозах, что подтверждено многочисленными исследованиями и не вызывает привыкания.
Возможности соматотропина в реювенилизации
Другим подозреваемым на роль цитокинина мог бы быть гормон роста – соматотропин (СТГ). Выделяется наиболее интенсивно в детском и подростковом возрасте, и его уровень постепенно снижается по мере старения. Однако, даже после достижения взрослого возраста, этот гормон продолжает оказывать влияние на многие процессы в организме. Гормон роста играет важную роль в поддержании здоровья и восстановлении тканей, улучшении общей физической формы, а также в омоложении организма. Но, тем не менее, его правильней относить к аналогу у растений – гиббереллину. Введение его в завышенных дозах у людей пожилых не делает «чудеса», как это смог бы цитокинин у стареющих растений, вплоть до регенерации, закладки новых ростовых точек в стареющих онтостадийно побегах. Реального омоложения от СТГ нет.
Существуют ли неизвестные факторы реювенилизации аналоги цитокинину?
Подозреваемыми могли бы стать микроРНК, роль которых пока не до конца изучена и осмыслена. Напомню, что цитокинин способен запустить регенерацию, полную реювенилизацию, снятие онтостадийности и стать хорошим маркером старения, так как он способен отменить симптомы старения на желтеющих листьях и отмену их апоптоза. У человека таким маркером могло бы стать вещество, которое отменяет седину волос, явление которое является типичным и универсальным проявлением сенилита или сенесцента.
Очевидно угасание СТГ с возрастом является всего лишь проявлением гомойотопии, то есть принципа ограничения размеров животных, и на определенных стадиях развития рост должен быть остановлен. Со старением это никак не связано.
А может в организмах животных такого гормона аналога нет? Организм человека вполне способен справиться без него и без открытия резервов тотипотентности? Он вполне может существовать за счёт комплекса других гормонов?
Онтостадийные этапы развития-созревания гипоталамуса
Организм человека от первого дня своей жизни и до последнего дня проходит через пять четко выраженных этапов онтогенеза, каждый из которых имеет свой гормональный профиль. Их можно назвать этапы онтогенеза. Это следующие: детство, пубертат, юность, адальтус, матура и сенилит. Точно также такие же изменения происходят и в гипоталамусе. Он тоже проходит через эти этапы. Каждый этап – это новая стадия зрелости организма и гипоталамуса, то есть, по другому, онтостадийные продвижения. Такая непрерывная динамика изменений представляет элевационный процесс. Организм и гипоталамус проходят через ряд ступеней развития, движутся по ним как по эскалатору. Каждая ступень развития имеет свои параметры и программы для включения на генетическом уровне.
Развитие гипоталамуса происходит одновременно со всем организмом; одно подготавливает почву для другого, но и в тоже время гипоталамус является ведущим механизмом. Организм проходит через ряд онтогенетических фаз развития, а вместе с ним и гипоталамус.
Но следует понимать, что ювенильная сома имеет постоянно открытую программу и драйвера на непрерывный рост, а он всегда имеет предел, за которым его надо тормозить. Начинаются проблемы гомойотопных* организмов, когда организм постоянно усиливает, наращивает тормозные процессы из гипоталамуса. Поскольку он матрикален*, то должен работать на принципах непрерывного наращивания тормозных процессов.
Как видим, тип онтогенеза может задаваться как с клеточного нижнего уровня, так и с верхнего гипоталамического или апикального.
Гипоталамус животных можно назвать прибором многоразового пользования, но который тоже имеет свои пределы. Закрытие возможностей гипоталамуса происходит постепенно и растянуто во времени, оно же определяет его возможности по руководству половозрелой системой организма, темпы его постепенного созревания и перезревания. Такое перезревание аналогично тому, что происходит при гипердоминантном генеративном созревании у монокарпических растений.
С возрастом на фазе перехода на этапы онтогенеза матуру, адальтус и сенилит состояние гипоталамуса определяет принцип стем-соматического или прогениторного доминирования, когда исчерпаны возможности стволовых клеток, а доминируют соматические и концевые типы стволовых (прогениторные), с минимальной степенью потентности к репарациям, которые не позволяют дальнейшему самовосстановлению его, например как это происходит у растения алоэ древовидного.
Вывод. В человеческом организме происходит непрерывное, но замедленное перезревание гипоталамуса. Такое перезревание по сути является принципом гипоталамического доминирования, что аналогично апикальному доминированию. Апикальное доминирование может перейти в гиперапикальное доминирование, что является причиной монокарпизма и жесткого лимитирования онтогенеза. В человеческом организме гипергипоталамическое доминирование (ГГД) тоже является лимитирующим фактором его онтогенеза. Гипоталамус также как и апекс быстрее всех набирает зрелость, переподчиняет себе ткани-мишени, а затем он же быстрее других структур проявляет все симптомы старения. Он орган-водитель старения = сенилита. Он проявляет старение, но не является его первопричиной. Будучи органом-водителем он одновременно и сам является ведомым. Неудивительно, что именно в нём преобладает накопление провоспалительных веществ. Ткани с провоспалением отличаются тем, что выделяют повышенное количество сигнального белка — так называемый фактор NF-kB, который включает клеточную реакцию на самовоспаление. Выявлено, что самая значительная концентрация белка NF-kB отмечалась именно в гипоталамусе. Клеточное провоспаление – это важнейшее проявление сенесцента, когда клетки теряют свою функциональную работоспособность. Ряд авторов утверждает, что клеточное старение первично и это основа системного воспаления, которое запускает общее старение. Это противоречит и отрицается многочисленными приведёнными данными в нашей теории. Таким образом, гипоталамус проявляет двойственность в старении, а точнее в геронтосе: изменения в нём первично происходят на уровне сенилита, а затем на уровне сенесцента. В связи с подавлением, дегенерацией эпифиза происходит разблокировка гипоталамуса, генеративная зрелость которого лавинообразно увеличивается по экспоненте, заходит за пределы оптимума, выходит за рамки коридора гормезиса, но затем также лавинообразно разрушается, дегенерирует в виде провоспаления и заходит в сенилит с усыханием, увяданием трофических функций. Внешние проявления такого трофического увядания хорошо отображены на рис. 31 с одной и той же женщиной в возрасте 18 и 118 лет.
Попутно отмечу, что мелатонин – гормон эпифиза - имеет важнейшее свойство – противодействовать провоспалению, но чем выше возраст, тем меньше мелатонина вырабатывает эпифиз. Задача найти способ эндогенного повышения выработки мелатонина в эпифизе или решить проблему экзогенного введения мелатонина.
Если нам удастся найти возможность преодоления этого принципа Гипергипоталамического доминирования, когда происходит угасание в очагах стволовых клеток многочисленных тканей, станет возможным обойти лимитирующий онтогенез фактор, который обуславливает гипоталамус. Но это не значит, что для этого надо воздействовать именно напрямую на гипоталамус, правильнее идти по пути активации стволовых клеток во всём организме с целью выйти на путь сомато-стемного доминирования.

Рис. 31. На рисунке приведены фото одного и того же человека через 100 лет, чтобы показать особенности увядания, угасания человеческого организма.
Снять, отменить эффект подавляющего действия гипоталамуса можно в условиях, когда ему будет более сильное противодействие в виде Жизненной Силы = Витаукт со стороны всей массы стволовых клеток.

Рис. 32. Агава – растение, которое цветёт только один раз в жизни = монокарпик с гиперапикальным доминированием.

Рис. 33. Древовидное алое может цвести многократно.
Гипоталамус можно представить как систему трансформера, которая непрерывно разворачивается, раскрывается и проявляет свои новые возможности и состояния: из незрелой он переходит в зрелую, а затем перезрелую фазу развития. Динамической силой его разворачивания в детстве и юности является непрерывный рост на периферии. Трансформинг начинается еще во внутриутробном периоде развития, проходит через все этапы, ступени онтогенеза и заканчивается на этапе инволюции, угасания. Трансформинг это по сути созревание, становление органа, появление на нём рецепторов. Позже созревают рецепторы на органах-мишенях, налаживаются связи «верха» и «низа», выстраивается соподчинённость. По сути, это так называемое ступенчатое становление, когда становление новой фазы развития не возможно без завершения становления предыдущей.
В последующем, когда заканчивается юность и останавливается рост, динамической силой дальнейших перестроек в нём являются не количественные перестройки, а качественны, когда уменьшается количество гормона роста и мелатонина, но увеличивается половая зрелость, количество гормонов мужской и женской сферы максимально высокое. Это ведёт к матуре – сверхзрелости, а матура ведёт к перезрелости – адальтусу. Адальтус это состояние далеко за пределами оптимума и гормезиса, когда сил Витаукта недостаточно, поэтому он неизбежно кренит к климаксу и сенилиту.
Гипоталамический синдром – неминуемое проявление возрастных перестроек и сенесцента
Выше мы обсуждали о возрастном воспалении гипоталамуса, как одной из причин гипоталамического синдрома. Любая ткань, что стареет, несёт в себе элементы провоспаления. Причем старение на уровне тканей многократно усиливается на уровне гипоталамуса. Гипоталамус стареет не сам по себе, а в ходе старения всего организма. Провоспаление в области гипоталамуса – одна из причин нарастания его резистентности, нечувствительности, толерантности его центров регулировок к корректирующим сигналам. Причём перестройки активности гипоталамических центров происходят ассиметрично, какие-то ядра усиливают свою активность, другие её ослабляют. Гипоталамус человека относится к структуре способной обеспечивать долгожительство, он и сам определяет темпы старения, но и зависит от старения организма в целом. Он определяет сроки климакса, следовательно является драйверным звеном в разворачивании онтогенеза и старения. Он задаёт темпы старения организму, но при этом он же является и буфером сдерживающим старение. Казалось бы это утверждение противоречит тезису о том, что старение на уровне тканей многократно усиливается на уровне гипоталамуса. Да, действительно, у животных с сильно развитым гипоталамусом, проработанными ядрами, ведущими к балансу между центром и периферией, гипоталамус может на многие годы и десятилетия отодвинуть старение, но когда потенциал старения накапливается на уровне сомы-тканей до критической линии, темпы старения в гипоталамусе начинают обгонять темпы старения во всём организме. Здесь происходит временное подобие ауксиновой молодости, но нет подобия подлинному цитокининовому омоложению.
Так что старение организма происходит одновременно на всех этажах организма. Воспаление – это один из универсальных признаков старения, не взирая на то с какого направления оно зарождается – клеточного или организменного. Очень важно разобраться в том, что является первопричиной гипоталамического синдрома – уровень клеточного старения сенесцента его или уровень сенилита, или это одновременный процесс с двух этажей?
Подсказкой является эксперимент на мышах с их парабиозом, когда сшивали кровеносную систему молодой и старой мыши и получали омоложение старых. Если бы парабиоз не смог омолодить гипоталамус не возможно было бы достигнуть омоложение. Следовательно, снизу за счёт изменения активности стволовых клеток и их стимулирования происходит одновременно и процесс омоложения гипоталамуса. Очевидно, существуют универсальные вещества молодости, которые влияют одинаково на любые типы тканей, а также на нижние и верхние этажи организма.
Итак, гипоталамус мы сравнили с деятельностью матрикального апекса деревьев. Но у каждого дерева есть множество апексов. Они находятся в камбии и на почках ветвей I, II, III, IV и т. д. порядков. Всю их деятельность можно сравнить со стволовыми клетками различных тканей в организме. Их задача обновлять и восстанавливать, регенерировать. Только у растений это происходит ростом наружу, а в животном мире это происходит за счет обновления в очагах роста старых клеток на новые (точки самообновления).
Принцип гомойотопии – особенность существования с ограниченными размерами в мире животных
Причём следует учитывать, что особенностью современных животных является необходимость проявления у них гомойотопии, то есть сохранения принципа постоянства размеров. Если у растений вместо старых веток появляются новые, молодые на новом месте, то у животных это должно происходить строго внутри, не выходя за размеры данной особи, то есть обязательна аутозамена старых на новые или усиление репаративных способностей у клеток не подлежащих замене (нейроны и др.).
Рис. 34. Расположение разнокачественных ветвей и побегов в кроне. На кончиках этих ветвей находятся в зависимости от порядка ветвления онтостадийно разнокачественные почки. По сути, они являются аналогами стволовых клеток в различных тканях организма животного.
Ослабление механизма аутосенолитизма – одна из важных причин старения организма
Такая система замены клеток («точки самообновления») у животных намного сложнее регулируется и усложняется тем, что должны быть созданы механизмы устранения старых линий клеток на более молодые. То есть должны работать непрерывно пейсмекерные (на основе чередования ритмов) механизмы самоомоложения, самовосстановления с условием непрерывного устранения старых клеток. Очевидно, для этого должны быть отработаны специфические механизмы самоликвидации стареющих линий клеток с помощью аутосенолитизма (самоуничтожение стареющих клеток), которые реализуются с помощью механизмов апоптоза, аутофагии и аутолиза. Проблема в том, что стареющие линии клеток (приобретшие сенесцентность) чаще всего сами не отмирают и при этом перекрывают дорогу для появления молодых, полноценных. Точка самообновления не работает.
Сенолитизм (механизм самоустранения старых линий клеток) – очевидно этот механизм проработан у животных ограничено и не проявлен до конца. Очевидно, это нужно с целью дать возможность филогенезу не ограничивать эволюцию и в то же время оптимизировать долгожительство, управляя им с помощью отбора?
В тоже время в ряде работ показано, что процесс старения связан с утратой клеткой способности избавляться от поврежденных компонентов и погибших патогенов. Повреждение или дисфункция лизосом является отличительной чертой процесса старения и развития многих заболеваний, в первую очередь, нейродегенеративных. Для реакции лизосомального повреждения необходим белок под названием серин-треонинкиназа 38 (STK38).
Парабиогенное питание или о роли корешков проростков растений для преодоления сенесцента (клеточного старения)
О целебной силе пророщенных зелёных ростков сказано много, но при этом нет никакой информации о значимости именно их корневой части как источника реювенилизирующих цитокининов, а также всего комплекса оживляющих весь организм ферментов, внутриклеточных гормонов, факторов роста, РНК. Цитокинины обладают способностью противодействовать гормону стресса и старения – абсцизовой кислоте, а также ингибиторам, гормонам покоя. Они вырабатываются в апексах (ростовых верхушках) корней и поддерживают существование мезенхимных = эмбриональных тканей растений даже во взрослом их состоянии. Именно от мощи и способности самовосстановления корней зависит продолжительность жизни того или иного вида растений, определяет быть ли ему 1-, 2-х или многолетним растением, вплоть до тысячелетий, как например Тополь осинолистный, вегетативная роща которого существует 80 тысяч лет [2]. Вдумайтесь, от 1-го до 80000 лет.
Такой гормональной ювенилизирующей непрерывной поддержки из корней по увеличению степени клеточной потентности как у растений, дающая им возможность неограниченного самовозобновления, в организме животных нет. Кроме того, и стволовые клетки I-го порядка отсутствуют, а возможность программативного порядка возврата на этот уровень закрыта. Это резко отличает клетки животного типа от клеток растительного типа, у которых имеются специальные триггерные программы по переключению степени потентности и возврата на клетки I-го порядка. Поэтому стволовые клетки животных с более высоким порядком потентности с возрастом «глохнут». Стволовым клеткам II и III-го порядков потентности нужна поддержка. Только клетки I-го порядка, то есть тотипотентные, в этом не нуждаются. Очевидно, это является ограничительной особенностью клеток животного типа, что и проявляется только у них в виде предела Хейфлика* и сенесцента*, а также онтофизиса*.
Конечно, цитокининов (гормона возобновления и молодости) в организме животных нет, но автор считает, что назначение корней растений не просто снабжать их нутриентами, но не менее важной задачей – пробудить и обеспечивать энергией роста, Жизненной Силой верхнюю часть растения. Эта же Жизненная Сила юных корешков также может оказать пробуждающее действие как на сенесцентные клетки, так и активирование стволовых у человека. В данном случае ожидается, что приём супрафизиологических доз, то есть завышенных многократно раз выше потребности организма в этих веществах, гомогената из таких корешков окажет не гормональное действие, а за счёт парабиогенеза* по «оживлению» клеток у человека, то есть всем клеточным секретомом, комплексом веществ стимулирующих рост клеток, а точнее точек самовозобновления. Нечто аналогичное мы описывали ранее в книге Гарбузова Г.А.: «Биология молодости» [2] по эффекту гетерохронического парабиоза, когда при сшивании кровеносной системы молодой особи мыши со старой происходило омоложение последней и увеличение продолжительности жизни. Но в нашем понимании молодые ростки (корешки) благодаря своей сверхмощной энергии роста оживляют и омолаживают клетки из другого типа живого - животные.
Гомология эффекта парабиогенеаза от приёма ювенильных корешков в супрафизиологических дозах реювенилизирующему эффекту на биотроне Ц. Каньжэна.
Можно предположить, что реювенилизирующее воздействие от гомогената из корневой части ростков будет происходить отчасти не только за счёт неких химических субстанций, ростовых факторов или другое, но и за счёт совершенно другого принципа, нечто подобного биорезонансному полю в биотроне доктора Цзяна Каньжэна, в котором как через линзу фокусировалось и многократно усиливалось биоэлектромагнитное поле данного живого объекта. При этом усиленное многократно поле этого объекта размещали рядом с другим и поле первого объекта меняло ряд биологических параметров другого. В данном случае усиленное поле (митогенетическое излучение Гурвича?) первого объекта было доминирующим и каким-то образом подстраивало чужие часоты себе в резонанс под себя таким образом, что через магнитные поля перенастраивало генопрограммы морфогенетических параметров другого так, что оно меняло некоторые формы на более подобные первому. Кроме того, метод проявил себя как средство реабилитации ДНК путём проведения воздействия с помощью биоэлектромагнитного поля молодых организмов на старые. Очевидно, это подсказка способа преодоления проблемы сенесцента. Обращает на себя внимание, что реализовать это было возможно только через посредство донора биополей взятого на эмбриональном этапе развития, то есть в активной фазе становления морфогенеза при прохождении через «трансформинг», разворачивание зародыша, на фоне активного проявления состояния максимальной клеточной ювенальности. В указанном методе описывается про возможность некоторых морфогенетических изменений, но для нас представляет бо¢льший интерес добиться онтогенетических изменений путём реювенилизации и усиления жизненной активности стволовых клеток, которые с возрастом «тухнут». При этом исследователь для целей омоложения и лечения использовал ростки, в частности пшеницы и другие, но активность поля их Жизненной Силы нужно было усилить многократно, для чего он применял свой биотрон. Ключевое слово: усиливал.

Рис. 35. Схематичное объяснение принципа действия биотрона.
Дело в том, что сенесцент клеток животного типа связан с торможением митотического цикла на фазе G0 (покоя), что ведёт к репликативному ступору (невозможность самообновления клеток за счёт непрерывных делений), что сопровождается депрессией клеточной энергии, в виде АТФ, митохондриальной и дифференциальной дисфункциями, эпигенетическим изменениям, электростатическим изменениям на хроматине и отключению за счёт этого программ рециклинга, то есть полного самовосстановления, в том числе и на уровне теломеразы и теломер. Все эти проблемы можно «пробить» либо соответствующими гормонами, либо парабиогенезом. Этот репликативный ступор сопряжен с эпигенетическими сдвигами. Если сенилит связан с гормональными сдвигами за пределы гормезиса, то сенесцент связан со сдвигами за пределы эпигенома баланса и выхода за пределы гомеостаза. Для достижения антисенесцентного эффекта нужно употребление довольно больших количеств корешковой массы. Это около 100-300 г в день в виде гомогената полученного на блендере из них. Возможно, чтобы еще более усилить эффект, необходимо будет из этого гомогената извлекать концентрированный сок. Для этих целей наиболее подходит двухшнековая соковыжималка Angel Juicer Angelia. Преимущества её в максимальной степени иссушения жмыха, работает на малых оборотах, не нагревает сок, не даёт ему окисляться на воздухе.
За основу лучше использовать для получения корешковой массы зёрна пшеницы, гречки, подсолнуха, чечевицы, горошка… Их проращивают обычно от 2-3 дней или 4-6 дней и несколько более до образования обилия нежных корешков, которые следует отделить от зерновки и верхнего ростка. Лучше для более высокой эффективности использовать преимущественно массу корешков, хотя можно использовать и всю биомассу проростка.
При проращивании следует соблюдать стерильность, для чего используют поддоны с двойным дном из сетки. Содержать поддоны лучше в специальных тёплых плёночных домиках. Корешки при этом проникают вниз через сетку и висят в воздухе, съёмный поддон двойного дна вытаскивают и соскабливают снизу корешки. Должна быть предусмотрена система регулярного автоматического опрыскивания или туманообразования. Все подозрительные или больные растения тщательно удаляют.

Рис. 36. На рисунке изображены хорошо развитая нежная волосковидная корневая система проростков пшеницы, которую и следует заготавливать отдельно и применять после размельчения в блендере.

Рис. 37. На фото видно бобовые проростки маша, как лучшие ингредиент для здорового питания. Перед употреблением лучше измельчать до пюре на блендере.

Рис. 38. Микрозелень люцерны. Иногда происходит путаница, когда семядоли на стволике путают с корешками. У семядолей всегда на конце образуется зачаток листочков.
Сенесцентные клетки-зомби — ложный след в расследовании сути старения?
Концепция первичной сенесцентности клеток приобретает всё большую весомость и доверительный уровень и переходит в разряд главенствующей парадигмы. От неё начинают отталкиваться и достраивать новые веточки концепций. Но, очевидно, здесь какая-то ошибка, неверная трактовка не позволяющая выйти на путь истины. Вспомним, что у растений клетки, по сути, бессмертны. Так выразительным примером может быть столоновая (корневыми отпрысками) «роща» Тополя осиноподобного, которая обладает особенностью вегетативного саморазмножения корневыми отводками и известен её возраст как одной линии организма существующей 80000 лет и не проявляющей никаких признаков клеточного старения и вырождения.

Рис. 39. Феномен одного супердерева Тополя осиноподобного в виде клональной колонии, напоминающей рощу, на площади 43 га и весом 6000 тонн чистой массы. Растение распространяется вегетативным путем корневыми отпрысками. Предположительно возраст этой рощи около 80000 лет.
Поэтому следует уметь объяснить причину отсутствия клеточного старения у растений и на основании этого дать правильное объяснение «сенесцентности» у животных. В последние годы популярно стало понятие сенесцентности*. В основе этого лежит лимит Хейфлика. При этом возникают некие клетки-зомби, которые мало того, что уже не выполняют свои функции, проявляют признаки нечувствительности к сигналингу, так ещё и отказываются вовремя умирать (апоптировать), уступая дорогу молодым полноценным клеткам, при этом ещё и отравляют всё вокруг себя, выделяя коктейль провоспалительных веществ, называемый “senescence-associated secretory phenotype” или SASP. Это можно расценить как бессмысленное пролонгированное существование сенесцентирующих клеток, которые и сами не работают, и другим не дают возможности. Эти клетки живут без поддерживающих их репараций, которые должны быть обеспечены механизмами аутофагии. Ни репарации, ни репликации. При этом клетки похоже «застывают», заблокированы на G0-фазе клеточного цикла, то есть покое. Таким клеткам нужна какая-то встряска, чтобы перезапустить их работу. Таковыми, очевидно, могут быть циклические периоды голода?
Концепция сенесцента – ложный след?
В норме блок G0 происходит при помощи белков — супрессоров митоза, один из которых — ретинобластомный белок (Rb-белок), кодируемый нормальными аллелями гена ретинобластомы. Данный белок прикрепляется к особым регуляторным протеинам, блокируя стимуляцию транскрипции генов, необходимых для пролиферации клеток.
То, что в организме по мере старения становится больше сенесцентных клеток, ещё не означает, что они являются драйвером старения. Имеются подозрения, что концепция сенесцентности может оказаться ложным следом.
Это поддерживают некоторые исследования.
Сенесцентность клеток – это всего лишь онтофизисная форма проявления сенилита
Описанные выше данные являются подсказкой того, что в организмах сенесцентность клеток есть, но зависит от сенилита, который и является первичным определяющим фактором.
Сенилит обуславливает ограничение функции эпифиза и снижению минимум в 2 раза уровня выработки мелатонина. Как результат усиление оксидатиного или клеточного стреса. Это в свою очередь ведёт к измнениям эпигенетическим и на хроматине, итогом чего является остановка работы теломеразы с укорочением и повреждением теломер, что в итоге ведёт к репликатиному торможению или сенесценту.
Феномен Хейфлика, очевидно, связан с тем, что клетки в культуре ткани отлучены от мелатонина, что в итоге ведёт к онтофизису и сенесценту.
Это результат «впечатывания» динамики онтогенеза не на уровне гормональных перестроек, и которые могут быть смещены, а на самом нижнем клеточном эпигенетическом уровне. По сути, сенесцент – это проявление онтофизиса. Онтофизис может быть следствием одной из закреплённых фаз онтостадийности и при определённых условиях он может происходить скачкообразно, перескакивая все предшествующие ступени онтостадийности. Именно таковым «перескоком» является и пример лимита Хейфлика, который является проявлением стресса для клеток и хроническим их провоспалительным статусом с итогом выхода на митохондриальную дисфункцию.
Исходя из приведенных развенчивающих данных и онтофизисной концепции сенесцента предложенной Г.А. Гарбузовым, лимит Хейфлика это всего лишь экспериментальный артефакт не правильно осмысленный? Ошибка трактовки «предела Хейфлика» очевидна: исследователи имели дело с изолированными клетками. Они забыли: отдельные клеточные культуры ткани животных так же не способны полноценно существовать, как и ампутированные органы. Клеточный уровень определяется жизнью организма в целом.
Взаимоотношения гипоталамуса со стволовыми клетками различных тканей
Выше было обсуждено сходство принципа работы гипоталамуса и апекса древесных растений, которые и определяют особенности фазы старения = сенилита организма. Апекс управляет возобновлением ткани не через соматические клетки, а через её стволовые клетки, угнетает или активирует их, ведёт к трансформациям в них. В свою очередь состояние стволовых клеток определяет состояние апекса. На начальных этапах развития, очевидно когда происходит рост, можно говорить о соматически-стволовом = стем доминировании, когда в организме поддерживается состояние молодости.
Стем-доминирование
Это когда преобладает становление, разворачивание и стимуляция с низу, а не с верху. С верху идёт только координирование-дирижирование ростом. У человека такое стем-доминирование продолжается до 12 лет, на этапе роста и становления. После этого фазы ювенильности и предпубертатности заканчиваются и начинается генеративный этап развития. Причём это состояние ювенильности не является простым отсутствием зрелости, готовности к генеративным проявлением, а активным подавлением, отодвижение наступления зрелости. У растений таким механизмом отодвижения является наличие повышенного уровня гормонов молодости цитокининов, у человека, очевидно, такими функциями обладают фетальные белки или микроРНК. Для ювенильности тоже нужна своя гормональная база, когда генеративный гормональный фон не является доминирующим. Но простое кастрирование животного, тем не менее, не решает проблему поддержания статуса ювенильности. Понятие стем-доминирование в биологии введено впервые Гарбузовым Г.А. Обычно стем-доминирование проявляет себя на фетальном (эмбриональном) и натальном (детство) этапах онтогенеза. Постепенно меняется на гипоталамическое доминирование с переходом в гипергипоталамическое.
Возрастной переход на гипергипоталамическое доминирование
Но с возрастом и старостью можно говорить о гипергипоталамическом доминировании, когда полностью остановлен рост и происходят качественные изменения адальтуса - зрелости. Количественные изменения заменяются на качественные. Причём фазы, когда наступает равновесие и остановка нарастаний дальнейших качественных изменений нет. Выход за рамки равновесия и гормезиса только увеличивается и доходит до критических величин.
Таким образом, онтогенез является движением (элевацией) от стем-доминирования к гипергипоталамическому доминированию. Ранее В. Дильман [8] утверждал о сугубо гипоталамическом элевационном процессе как движущей силы онтогенеза. Но это недопонимание всеобъемлемости процесса, который затрагивает все этажи организма.
В свою очередь гипоталамус у молодых также влияет на активность стволовых клеток в тканях и подстёгивает возобновления, омоложения в них. Чем лучше гипоталамус выполняет эту роль, тем более продолжительные этапы онтогенеза, тем более к группе долгожителей будет относится данный вид. Теоретически можно было бы рассчитывать, что при идеальной работе гипоталамуса жизнь данного вида могла бы быть не лимитированной. Но это возможно могло бы быть только на фазе юности. В биологии развития известно, что чем больше фаза ювенильности, тем выше долгожительство. Венчает эту линейку человек, детство и пубертатный период у которого самый длинный - до 12 лет.
Эпифизарное доминирование или эпифизарная молодость с гипервысоким уровнем мелатонина - гомолог радиксальномому доминированию у растений с гипервысоким уровнем цитокинина
Радиксальное доминирование аналог эпифизарному доминированию
Напомню что у растений фаза вегетативной молодости поддерживается гормоном корней цитокинином. В обоих случаях их гормоны цитокинин и мелатонин вырабатываются в темновых условиях. Оба они являются гормонами ювенильности (неполовозрелости) и вегетационной фазы онтогенеза (отсутствие плодоношения). Оба являются антагонистами генеративным гормонам как ауксины у растений и половые гормоны у животных, а также гормонам стресса (абсцизовая кислота у растений вызывает и старение) и кортизол у животных. Радиксальное доминирование в определённых формах онтогенеза позволяет реализации неограниченного принципа существования как у сосны остистой (4800 лет) и у тополя осинолистного (80 тысяч лет). Но человек не относится к группе с таким типом онтогенеза: выработка мелатонина в нём не поддерживается неограниченно и начинает гаснуть с 5 лет. Дальше руководство гормональным фоном полностью переподчиняется на гипоталамус, роль эпифиза пстепенно снижается, а гипоталамуса увеличивается. Снижение выработки мелатонина понижает степень чувствительности гипоталамуса к обратному подавляющему сигналингу. Гипоталамус становится работать на более высоких оборотах, более подобных функционально разболтанным, то есть работать за пределами оптимума для требований организму. Гипертрофическая функция эпифиза гаснет за ненадобностью, а остаётся только циркадная, что можно осуществлять на уровнях мелатонина в разы ниже. У молодых людей, а также лиц пожилого возраста признаки кальцификации обнаруживаются в 35-40 % случаев, а к 65 годам в 100% случаев эпифиз полностью перегружен известковыми отложениями. Возможно, у детей и людей среднего возраста этот кальциноз является прологом к преждевременному старению. Конечно, дети 5 или 10 летнего возраста не становятся стариками, их онтогенез развивается под общим руководством гипоталамического доминирования* и согласно его программам, но всё это ускорение и усиление со стороны гипоталамуса будет происходить на фоне угасания со стороны эпифиза. Так поседение или климакс могут начаться и в 30 и в 50 и более лет.
Что означает старение эпифиза?
Тем не менее, существуют научные обзоры, которые приводят несколько отличающиеся данные [11]. Так к 50-60 гг. жизни происходит увеличение размеров эпифиза с 80-100 мг до 150-160 мг - главным образом вследствие фиброза, который не связан с уменьшением числа паренхимальных клеток. В пинеалоцитах происходит возрастное уменьшение соотношения объемов ядра и цитоплазмы, указывающее на гипофункцию клеток. Сделан вывод, что у людей "истинной сенильной атрофии шишковидной железы не происходит". Патоморфологическое исследование эпифиза людей, в том числе старше 70 лет, приводит в заключению, что эпифизы стариков и молодых людей почти не различаются с помощью световой микроскопии. Электронная микроскопия показывает, что в пинеалоцитах при старении происходит увеличение числа липофусциновых гранул и инвагинаций ядра. Накопление липофусцина и фиброз характерны для возрастных изменений в самых разных тканях. Кроме липофусцина в эпифизе при старении накапливаются кальциевые конкреции, представляющие собой отложения гидроксиапатита на органическом ядре. У крыс при старении в эпифизе увеличивается как содержание кальция, так и число кальциевых конкреций, главным образом в более темных пинеалоцитах, имеющих признаки дегенерации. Доля тёмных пинеалоцитов с возрастом увеличивается, а общее число пинеалоцитов снижется - примерно на 18% к возрасту 28 мес. Накопление кальциевых отложений выявлено в набухших пинеалоцитах монгольских песчанок. Предполагают, что оно происходит из-за возрастного снижения активности Са++-АТФазы и ведет к смерти клеток.
Содержание кальция. В эпифизах людей, погибших в возрасте от 3 мес. до 65 лет, общий уровень кальция находится в прямой корреляции с возрастом и в обратной - с ночным и дневным уровнями мелатонина в эпифизе. Чем больше мелатонина, тем меньше кальция и меньше старение эпифиза. Исследования кальциевых конкреций в клетках эпифиза людей в возрасте от 2 суток до 86 лет показало, что эти конкреции вряд ли имеют отношение к возрасту, поскольку могут отсутствовать даже у очень старых людей, а образование конкреций в пинеалоцитах связано скорее с секреторной активностью, чем с атрофией клеток. Следовательно кальциноз не неизбежный процесс.
Депривация симпатических нервов идущих в эпифиз песчанок удалением верхних шейных ганглиев приводила к тому, что накопления кальциевых конкреций в эпифизе не было. Следовательно, существенный вклад в старение эпифиза оказывает симпатический отдел вегетативной нервной системы. Таким образом, симпатическая часть нервной системы при её возбуждении подавляет эпифиз и угнетает синтез мелатонина. Чем больше возраст, тем сильнее активирована симпатика, но слабеет парасимпатика. Очевидно, парасимпатика усиливает эпифиз. Меняется нейромедиаторный фон и степень медиации эпифиза и гипоталамуса. Симпатика работает за счёт адреналина и норадреналина, а парасимпатика - за счёт ацетилхолина. Эхо всех этих колебаний и фиксирует эпифиз, будучи в роли эпифизоскопа* или сканера. Все эти изменения медиации меняют взаимоотношения эпифиза и гипоталамуса, растормаживая последний, но угнетая первый.
Функциональная симпатическая депривация эпифиза развивается по мере старения. Концентрация β-адренергических рецепторов в эпифизе у крыс при старении снижается так же, как это происходит, например, в полосатом теле и мозжечке. Это означает, что чувствительность эпифиза к стрессовым медиаторам резко снижается, что означает резкое снижение противодействия со стороны эпифиза стрессу. Бесконтрольный уровень активности симпатики растёт, а за ней ухудшается трофика присущая ювенильности. Кроме того, в эпифизе, как и в других отделах мозга, с возрастом ослабевает способность увеличивать число адренорецепторов в ответ на адренергическую депривацию. При старении снижается не только чувствительность эпифиза к норадренергической стимуляции, но и сама стимуляция. В эпифизе людей пожилого возраста и пациентов с болезнью Альцгеймера аксоны от верхних шейных ганглиев имеют признаки дегенерации. Иммунореактивность на тирозингидроксилазу, маркерный фермент катехоламинергических аксонов (в данном случае норадреналинергических), в эпифизе 25-мес. крыс в 2 раза ниже, чем у 4-мес. После односторонней симпатической денервации она у старых крыс, как и у молодых, снижается еще на 50%, но, в отличие от того, что происходит у молодых крыс, она не восстанавливается коллатеральным спраутингом. Это может быть следствием недостаточности как факторов роста нервов в самом эпифизе, так и регенеративной способности нейронов, иннервирующих эпифиз. Очевидно, снижена активность стволовых клеток из-за ослабления способности к репликациям, то есть сенесцента. Первая возможность маловероятна ввиду того, что уровень фактора роста нервов и его иРНК в эпифизе крыс при старении не снижается.
В возрасте, когда происходит выраженное снижение ночных пиков секреции мелатонина, в эпифизе не наблюдаются выраженные в той же мере дегенеративные изменения (в отличие, например, от тимуса, где снижение продукции его факторов прямо связано с дегенерацией самого органа). Возрастные изменения в эпифизе имеют более функциональную, чем органическую природу, что делает вероятным восстановление функций эпифиза.
Существенный вклад в возрастную гипофункцию эпифиза вносят нарушения во внешних по отношению к эпифизу норадренергических системах стимуляции его секреторной активности. Это подтверждается данными о повышении уровня мелатонина в эпифизе старых крыс, которым вводили депренил (ингибитор моноаминооксидазы-А). Действие ингибиторов МАО-А на эпифиз in vitro связано с ингибированием разложения норадреналина. Особенно сильная подверженность возрастным изменениям вообще присуща катехоламинергическим системам головного мозга, поскольку катехоламины являются отличным субстратом свободнорадикального окисления.
Функциональные последствия накопления кальциевых конкреций в эпифизе не ясны. Предполагается, что их образование происходит при чрезмерной стимуляции пинеалоцитов, как это может быть при стрессе, и является способом забуферивать ионы кальция и предохранять пинеалоциты от чрезмерного накопления ионов кальция в цитоплазме клеток. У людей уровень кальциевых конкреций обратно коррелирует с уровнем экскреции метаболитов мелатонина. Объяснения этому могут быть двоякие.
Очевидно, что накопление конкреций в ткани когда-то должно достигать степени, создающей непреодолимые помехи ее функциям, но неизвестно, какая это степень и достигается ли она в реальной жизни. С другой стороны, независимо от собственного вклада кальциевых конкреций в нарушения функций эпифиза, их уровень, а также уровень липофусцина, можно рассматривать как показатели кумулятивной дозы полученных пинеалоцитами повреждающих факторов эндогенного происхождения (кальциевые конкреции - показатель дозы избытка кальция, а липофусцин - показатель экспозиции клеток действию свободных радикалов кислорода). Одним из следствий накопления повреждений в макромолекулах из-за действия эндогенных свободных радикалов считается возрастное снижение способности клеток к индукции синтеза белков. В эпифизе оно может проявляться в снижении концентрации β-адренергических рецепторов и потере способности компенсировать усилением синтеза этих рецепторов адренергическую депривацию (которая при старении фактически и происходит).
Прямые данные о том, как влияет на старение эпифиза применение антиоксидантов, отсутствуют. С ослаблением свободнорадикального повреждения тканей связывают геропротекторное действие ограничения калорийности питания. Снижение концентрации β-адренорецепторов в эпифизе и других органах крыс замедляется ограничением диеты по калорийности. При этом активность ключевого фермента синтеза мелатонина, индоламин-N-ацетилтрансферазы, в эпифизе и содержание мелатонина в крови подопытных старых крыс оказываются выше, чем в контроле. Но из результатов измерения суточной экскреции 6-сульфатоксимелатонина только у тех крыс, у которых при старении не развиваются явные патологические процессы, следует, что ограничение калорийности питания не влияет на суммарную суточную продукцию мелатонина и её снижение по мере старения.
Уровень β-адренорецепторов в эпифизе и полосатом теле старых крыс удалось повысить введением S-аденозил-L-метионина, кофактора метилирования фосфолипидов мембран, который приводит к снижению их микровязкости, повышающейся при старении. S-аденозил-L-метионин является еще и кофактором метилирования ацетилсеротонина до мелатонина, поэтому причинно-следственные отношения в этом эксперименте не очевидны.
Секреция мелатонина у старых макак-резусов повышалась под действием синтетического пептида эпиталона. Эпиталон вводили старым самкам (20-26 лет) внутримышечно в дозе 10 мкг на животное (около 0,2 мкг/кг) в течение 10 сут. При этом вечерний (21 ч.) уровень мелатонина, сниженный по сравнению с имевшимся у молодых животных (6-8 лет) в 2 раза, повышался в 3 раза и оказывался выше, чем у молодых обезьян, на которых введение эпиталона в этом отношении не действовало.
Выводы:
- Отдельные функции эпифиза старых животных восстанавливались ограничением калорийности питания, введением S-аденозил-L-метионина или ингибиторов моноаминооксидазы-А. Восстановление способности эпифиза старых макак резусов секретировать мелатонин наблюдалось при введение синтетического пептида эпиталона.
Главенствующая роль эпифиза в начальных этапах онтогенеза и эпифизарной молодости
Выше мы описывали один из типов онтогенеза, когда происходит ювенильное доминирование гормона корней цитокинина, ведущего к неограниченному поддержанию стадии ювенильности = неотении и переводу таких растений в категорию долгожителей. Это так называемое радиксальное доминирование* или радиксальный тип онтогенеза*. Очевидно, похожую роль в организме животных играет эпифиз с его гормоном мелатонином, да и группой других пептидов. Известна роль эпифиза в качестве водителя циркадных (суточных) ритмов, но практически не осмыслена его участие и значимость в регулировке онтостадийных переходов. На эмбриональной стадии, то есть до рождения мелатонин почти отсутствует.
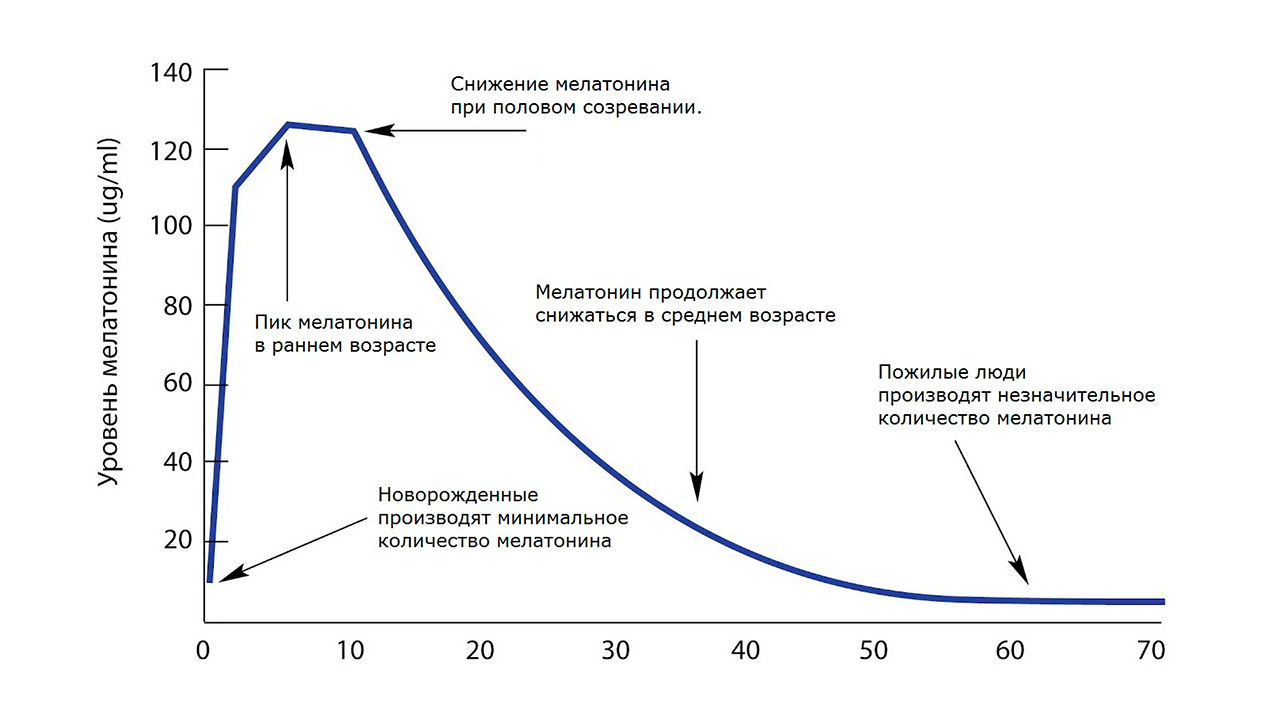
Рис. 40. На графике показана резко выраженная динамика подъёма уровня мелатонина десятикратно после рождения и как с наступлением половой зрелости его уровень резко снижается. Именно период увеличения его уровня определяет особенности этапа «эпифизарной молодости, тогда как снижение этого уровня определяет насупление новых фаз онтогенеза как юность, зрелость, перезрелость, климакс, старость.
Известно, что у человека максимальное его количество вырабатывается на натальной стадии развития (детства), а затем его уровень с 12 лет постепенно уменьшается и на стадии зрелости уменьшается десятикратно. У растений такая динамика взросления аналогична и совпадает с процессами утраты ювенильности, когда начинает срабатывать апикальное, или гиперапикальное доминирование и одновременно снижается и переподчиняется роль гормонов корней. Именно у молодых растений с переходом на новые этапы зрелости их корневое или радиксальное доминирование* ослабевает. По сути, апексы корней это антипод апексам вершин растений, которые регулируются другими гормонами. Но апексы радиксальные и матрикальные всегда сопряжены в едином круге, в одном случае доминируют радиксы, а в другом - матрикалы. И это является как динамической силой, драйвером стадий онтогенеза, так и может обеспечивать различные типы онтогенеза у различных видов. Также как и у корней функция эпифиза усиливается в темноте, корням нужна этиоляция, а эпифизу - темновой глубокий покой. Эпифиз подчиняется не только циркадным ритмам, но и сезонным, актиноритмическим и онтогенетическим. Чем выше возраст, тем больше его функция угасает.
Эпифиз можно приравнять к аналогу радиксального регулирования, а гипоталамус – к матрикальному = апикальному.
Эпифизарная молодость.
Только у человека хорошо развит этап эпифизарного доминирования. Он же совпадает с этапом эпифизарной молодости. По сути, это совпадает с расцветом Жизненной Силы и Витаукта* благодаря особенностям юношеского гормонального фона и мощи контроля над организмом со стороны эпифиза (уровень мелататонина в 2-3 раза выше чем в 23 года, а половых гормонов ниже). Этот период совпадает с максимальной обаятельностью облика и идеальным здоровьем (рис. 41), то есть максимальной устойчивостью ко всем патологическим факторам, что означает что все функции организма находятся в оптимальном русле гормезиса.

Рис. 41. На фото особенности фенотипа лица человека в период эпифизарной молодости. Он совпадает с максимальной обаятельностью облика и идеальным здоровьем, то есть максимальной устойчивостью ко всем патологическим факторам, когда все функции организма находятся в оптимальном русле гормезиса.
Эпифиз в роли «третьего глаза».
Долгое время эпифизу приписывали это мистическое свойство. Тем не менее, это определение в определённой степени соответствует действительности. Но только несколько в ином значении.
Эпифиз в роли камертона, локатора для циркадных внешних сигналов и в роли зонда, гироскопа онтогенеза* для внутреннего этапного саморазвития.
Очевидно, правильнее так его позиционировать за его способность анализировать не только внешнюю ситуацию, но и внутреннюю. Контроль суточных циркадных ритмов - это не единственное его важное свойство. Особый интерес представляет значимость его в роли онтостадийного регулятора. Нечто подобное гироскопу, в авиации это прибор, способный указывать и реагировать корректировкой на отклонение от заданных параметров курса; но в нашем случае имеется в виду механизм, который ориентирует организм о ходе динамики онтогенеза и при достижении определённых параметров отклонения включающий корректирующий сигнал на возрастные перестройки в гипоталамусе, то есть переход на следующую онтостадию. При этом в случае разблокировки гипоталамуса происходит обратное подавление работы эпифиза. Это в свою очередь отключает программу эпифизарной молодости. Он является «пусковым крючком» для запуска перехода на новую ступень. Только если гироскоп самолёта реагирует на отклонение угла от курса, то эпифиз реагирует на отклонение гормонального фона. Эту функцию эпифиза можно обозначить как эпифизоскоп, эпифизоскопию*. Основным его важнейшим свойством является способность проявлять эпифизарную молодость, расцвет которой продлевается до 12 лет и обеспечивается эпифизарным доминированием гормонального фона. Затем 3-5 лет наступает период паритета эпифизарного и гипоталамического доминирования. Это так называемый период раннего пубертата или молодости. В последующем наступает период адальтуса и смены внешнего фенотипа, аналогично тому как это происходит, например у термитов с резко выраженными отличиями.
Эпифиз в роли синхронизатора и гармонизатора.
К основной функции пинеальной железы относят синхронизацию биологических ритмов и поддержание нормального гормонального фона всего организма. Всё это связано с регуляцией циркадных ритмов.
Эпифиз в роли трофического фактора.
Очевидно, это еще одна его функция не до конца изученная и осмысленная. Возможно, тот статус гормонального состояния, который он поддерживает, или прямое воздействие мелатонина на ткани обеспечивает особый статус здоровья тканей, в том числе и высшее качество здоровья кожи. Мелатонин сдерживает уровень воспалительных веществ, в частности интерлейкина IL-17, который определяет фенотип старой кожи. Именно всё множество гормонов провоспаления (специфичных для каждого типа тканей) подавляют их трофику и являются базой для анизомалий, которые в них могут проявиться.
Эпифиз в роли зонда состояния вегетативной нервной системы (ВНС).
Эпифиз мощно иннервируется вегетатикой, а также сам сканируется и контролируется ею. Эпифиз это древняя система регулировки, а вегетатика – эволюционно продвинутая и вносящая свои коррективы в режим эпифиза, вплоть до смещения его значимости на второй план, подминая его главенство. Корректировка эпифиза происходит как иннервационно (по синаптическим каналам), так и за счет медиации – уровня медиаторов; тогда как гипоталамус корректируется до определённого уровня состороны эпифиза, но затем она ослабевает но переподчиняется на систему обратной связи с гормонами желез-мишеней. Так, депривация симпатических нервов идущих в эпифиз песчанок удалением верхних шейных ганглиев приводила к тому, что накопления кальциевых конкреций в эпифизе не было. Следовательно, существенный вклад в старение эпифиза оказывает симпатический отдел вегетативной нервной системы. Таким образом, симпатическая часть нервной системы при её возбуждении подавляет эпифиз и угнетает синтез мелатонина. Чем больше возраст, тем сильнее активирована симпатика, но слабеет парасимпатика. Очевидно, парасимпатика усиливает эпифиз. Меняется нейромедиаторный фон и степень медиации эпифиза и гипоталамуса. Симпатика работает за счёт адреналина и норадреналина, а парасимпатика - за счёт ацетилхолина. Эхо всех этих колебаний и фиксирует эпифиз, будучи в роли эпифизоскопа* или сканера. Все эти изменения медиации меняют взаимоотношения эпифиза и гипоталамуса, растормаживая последний, но угнетая первый.
Со старением вегетатики стареет и эпифиз? Так ли это? Может быть с изменением маятниковых рычагов ВНС происходит перекос в балансе симпатики и парасимпатики, что и угнетает эпифизис, а точнее происходит автодепривация эпифиза? Одновременно происходит снижение чувствительности в гипоталамусе, повышается его резистентность.
Автодепривация эпифиза – путь к гипоталамическому гипердоминированию и сенелиту.
Очевидно, последовательность событий созревания с выходом на адальтус и сенилит на примере песчанок следующая. Бурная половая активность песчанок и свет ведут к активизации симпатического отдела ВНС, а это угнетает работу эпифизу. С другой стороны активируется, растормаживается и непрерывно растёт работа гипоталамуса, что ведёт к гипергонадизму, а это дополнительно угнетает эпифиз, а также статус эпифизарной молодости. Таким образом, подавление эпифиза идёт в две фазы, что означает реализацию двойного фактора подавления эпифиза. Внешне эти различные фазы подавления эпифиза имеют различия и в фенотипах (адальтус и матура). По ряду данных именно двукратное снижение уровня выработки мелатонина является условием перехода со стадии онтогенеза адальтуса в стадию матуры (климакса), а за ним уже дорога к сенилиту. Очевидно, переходу на сенилит будет способствовать накопление сенесцентности, сенесцентиального состояния функционирующих тканей. Это уже будет третья ступень подавления эпифиза, которую следует обозначить как сенесцентную. Насильное угнетение, подавление пинеальной железы ведёт к воспалению и набуханию её клеток с последующим накоплением «мозгового песка», кальцификацией. А это и есть механизм автодепривации эпифиза. Снижение освещенности и ограничение половой активности растормаживают эпифиз. Выше мы уже обсуждали, что инволюция гипоталамуса сопровождается тем, что именно в нём максимально проявляются воспалительные процессы. Очевидно, такой процесс идёт и в эпифизе. Старение гипоталамуса идёт в две фазы: элевация и инволюция. Точно также динамика идёт и в эпифизе. А в целом это единый блок регулировок онтогенеза. Только, очевидно, в эпифизе идёт более глубокая инволюция с кальцинозом и более высокой степени утраты функциональности. Первым инволюционирует и выходит из строя эпифиз.
Пересадка эпифиза в вилочковую железу или область эпифиза старых мышей эпифиза от молодых приводила к увеличению продолжительности жизни животных и оказывала омолаживающее действие: изменялся их внешний вид и поведение. При пересадке шишковидной железы от старых животных молодым давало обратный эффект. Другие исследователи отметили, что ежевечернее питье воды с мелатонином на 20% удлиняет жизнь подопытных мышей и крыс. Возможно, это является конкретизацией феномена парабиоза, когда сшивали кровеносную систему молодой и старой мыши и получали омоложение у старой.
В чём отличие омолаживающих результатов пересадки эпифиза, гипоталамуса и половых желёз по Броун-Секару от молодых животных старым?
Следует отметить, что организм начинает стареть не снизу, а сверху. Следовательно, пересадка эпифиза даст более радикальный и устойчивый результат. Но эпифиз при этом тоже подзависмый орган от гипоталамуса и он тоже инволютирует. В связи с инволютивной резистентностью, нечувствительностью гипоталамуса можно ожидать, что «разбудить» его на фазе инволюции можно только более высокими дозами гормонов эпифиза. Простая подсадка гонад даст простой всплеск временной активации половой сферы и трофики тканей зависимых от них тканей. Подсадка части гипоталамуса тоже даст временный, но более выраженный эффект. Подсадка эпифиза, очевидно, даст более выраженный эффект, так как мелатонин имеет плюрипотентные свойства не только на половую сферу, но и на многие другие системы и железы. Но эффект опять же будет временным, поскольку перезревший гипоталамус и железы-мишени начнут его подавлять заново с еще большей силой. Эффект реювенилизации неминуемо постепенно сойдёт на нет.
Квазиэпифизарное доминирование
Задача научиться поддерживать состояние квазиэпифизарного доминирования. Это понятие отличается от понятия эпифизарного доминирования, которое сопряжено с циркадными ритмами и присуще юности - этапу онтогенеза. Квазиэпифизарное доминирование* – это состояние персистирующего принципа функционировния эпифиза с превалированием гормонов эпифиза не менее чем в 2 раза от нормы среднего возраста, когда полностью исключена возможность эпифизарно-гипоталамического паритета или переход на принцип гипоталамического доминирования*. Но вообще надо знать, что в ночной период уровень мелатонина в 10 раз >, чем днём. Следовательно организм легко переносит 10-кратные всплески. Квазиэпифизарное доминирование как раз и проявилось в случае летаргического сна с Одри Санто, который описан в этой книге. Следует отметить, что циркадность эпифизу придают супрахиазматические ядра гипоталамуса и ВНС, а переход на гипоталамическое доминирование обеспечивается за счёт открытой, неограниченной фазы роста функции гонад и гормона роста. Простая кастрация даже в детском возрасте тоже не приведёт существенно к длительному желаемому эффекту.
Циркадианная зависимость эпифиза.
Кастрация притормозит, но не остановит полную элевацию гипоталамуса, а с ним девиацию эпифиза, так как он еще связан с циркадианными функциями. Похоже это связано с тем, что существуют ритмы колебания работы эпифиза связанные с внешними факторами (циркадные) и с внутренними (циркадианные). Циркадный ритм – 24 часа, а циркадианный – 24 ÷ 28 часов. Циркадный ритм можно поменять, а циркадианный (врождённый) – нет. Эпифиз животных живущих под землёй переходит на циркадианный режим работы. Поэтому зрелость гонад у них не отменяется. За циркадианный ритм отвечает 2 супрахиазматических ядра, которые отвечают за хронометраж. Хронометрирование можно отключить только в летаргии, непробудном сне, как у Одри Санто? Если бы нам удалось сбить этот циркадианный ритм до 0 ÷ 3 часов, то можно было бы вернуть персистирующий статус эпифиза, то есть непрерывно вырабатывающий мелатонин. Есть животные, которые ведут подземный образ жизни, то есть темновой актиноритм = 24 часа, но, тем не менее, длительность жизни это им значительно не добавило. Исскусственное соддержание людей в длительной темноте (неделями) в условиях эксперимента без связи с внешним миром и контроля часов приводило к изменению циркадианного ритма с 24 ч до 25 часов. Организм автоматически хронометрирует циклы бодрствования и сна. Как добиться в лечебно-оздоровительных целях этого цикла до 0 ÷ 3 часов бодрствования и 21 ÷ 23 часа сна? Ведь именно во сне выработка мелатонина в 10 раз выше чем в фазе бодрствования.
Существует, очевидно, какой-то механизм регулировки, который «отпускает», освобождает эпифиз от циркадианного влияния и позволяет ему проперсистировать в определённом режиме, то есть выработать мелатонин. Обычно это происходит в глубоком сне с 24 часов ночи до 3 часов утра. Эта «свобода» персистировать отключается циркадианно. Очевидно, этот механизм находится в гипоталамусе и отвечает за засыпание или просыпание. Антифазой покоя в эпифизе могут быть уровень норадреналина и кортизола, то есть сигнал на возбуждение идёт как с надпочечников (кортизол), так и с супрахиазматических ядер, а также ядер где вырабатывает орексин. Блокировка этих путей приведет к удлинению ночной фазы сна.
Является ли зимняя спячка или анабиоз (гибернация), например у медведей, аналогом комы, летаргии или нарколепсии? Эта спячка может длиться до полугода. Пульс снижается с 55 ударов в минуту до примерно 9, а уровень метаболизма снижается на 53%. Естественно, все это время медведи не едят, не пьют и не производят отходы жизнедеятельности. Как им это удается? В организме, пребывающем в состоянии покоя в виде спячки, интенсивность всех обменных процессов сведены до физиологического минимума. То есть, энергии затрачивается ровно столько, сколько ее необходимо для того, чтобы зверь оставался живым и чтобы не произошли дегенеративные процессы в тканях и органах в связи с нехваткой энергии. Важно, что после 6 месяцев спячки и отсутствия физической активности у медведей не наблюдается ни одного из неблагоприятных последствий малоподвижного образа жизни у людей, таких как деградация мышц, сердечно-сосудистые заболевания, диабет 2 типа и почечная недостаточность. Очевидно, накапливается некий белок, который, останавливает процессы, ведущие к деградации и атрофии мышечной ткани.
Что такое нарколепсия?
В случае повреждения гипоталамуса происходит состояние нарколепсии, неограниченного сна и эпифиз переходит с циркадного на режим персистирования. Возможно у Одри Санто был реализован механизм полной нарколепсии?
Это заболевание нервной системы относится к гиперсомниям и характеризуется дневными приступами непреодолимой сонливости с приступами внезапного засыпания, приступами катаплексии, то есть внезапной утраты мышечного тонуса при ясном сознании, нарушениями ночного сна. Иногда отмечается кратковременная парализованность тела сразу после пробуждения. Открыт пептид, способствующий пробуждению. Следовательно существует центр ответственный за его выработку, а в случае его повреждения возможна ситуация нарушения ритма сна с её крайним проявлением в виде летаргии? Установлено, что дефицит гипокретина (орексина) является причиной нарколепсии. При нарколепсии гибнут нейроны вырабатывающие орексин (очевидно по аутоиммунной причине). В этом состоянии нарколепсии, очевидно, растормаживаются и механизмы непрерывной выработки мелатонина.
Механизмы циркадианности.
Орексин обеспечивает, проявляет циркадианность, но тормозит персистирование. Он накапливается днём, а мелатонин – ночью. Но орексин способствует просыпанию. Очевидно существует и вещество способтвующее засыпанию. Такой возможностью, очевидно, обладают тормозящий нейромедиатор — гамма-аминомасляная кислота (ГАМК) и аденозин? Не они ли запускают механизм персистирования и отключают орексин? ГАМК так же как и галанин, и мелатонин являются главным медиатороми сна. Но кто из них первичен, а кто вторичен? Очевидно, мелатонин всего лишь исполнитель, реализатор чужих поручений. Есть основания предполагать, что то вещество, которое запускает мелатонин, способно регулировать и его персистирование при условии его доминирующего статуса. Очевидно, в принципе достаточно запустить механизм персистирования мелатонина на 21 час и 3-х часовой циркадный режим, то этого достаточно будет чтобы удерживать состояние юности. Конечно, это не означает, что человек должен находится 21 час в состоянии сна, нет! Просто в состоянии бодрствования будет продолжаться выработка мелатонина.
В норме механизмы засыпания-просыпания должны быть абсолютно конгруэнтны, то есть подлажены друг под друга так, что происходит ритмичное лёгкое засыпание и лёгкое просыпание, для этого имеются два пртивоположных механизма, увязанных системой обратных связей в единый целый механизм. С одной стороны это орексиновый регулировщик, а с другой - ГАМК? Это два противоположных рычага единой маятниковой системы. Крылья этого маятника должны быть равны. Но что будет если это равенство будет смещено? Маятник просто не сможет работать, его заклинит. Это означает что маятник можно отключить с уклоном на стороне одного из рычагов. Появится состояние непробудного сна или хронической бессонницы. В истории известны случаи людей, которые были совершенно лишены сна и при этом себя чувствовали абсолютно нормально. В любом случае это выведет из строя механизм циркадианности и запустит принцип непрерывного персистирования или его полного отсутствия. В превом случае это будет квазиэпифизарное доминирование, а во втором - к отключению функции эпифиза, что в одном случае запустит механизм непрерывной автоювенилизации и неотении, а в другом приведёт к прогерии – преждевременному старению.
Снятие автодепривации эпифиза - путь к неотении?
Это означает, что для этого нужно превалирование парасимпатического крыла ВНС с его ацетилхолиновой медиацией?
Эпифиз координатор перехода от детства к юности и от юности к зрелости.
Из сказанного следует, что эпифиз держит под контролем динамику онтогенеза от эпифизарного доминирования до эпифизарной автодепривации. По сути, это путь от эпифизарного утра, эпифизарной молодости до эпифизарного вечера, заката или эпифизарной девиации* (отклонение от оптимального уровня активности, старости). Это означает выход за рамки оптимума жизнедеятельности организма, то есть за пределами гормезиса, что и определяет неизбежно мортальный исход.
Именно эпифиз определяет гипоталамусу время его элевации, чтобы перейти от стадии детства (постнатальный период) к стадии юности («имаго», с 5 до 12 лет, то есть уже не ребенок, но сформировавшийся подросток без признаков половозрелости, пубертата). В дальнейшем эпифиз определяет время перехода от юношества (начальный пубертат 12-15 лет) к молодости (половозрелости, полный пубертат, 16-20 лет), а затем к адальтусу. Раньше все эти функции приписывали к обязанностям гипоталамуса. Но при этом не учитывали, что гипоталамус лишь реализатор, партитура, подчиняемая более общим командам. Напомню, что удаление эпифиза в детстве разблокирует гипоталамус и ведёт его к досрочному созреванию и элевации, а на уровне желез-мишеней к гипергонадизму. Всё это регулируется соотношением уровней мелатонина и половых гормонов.
- Мелат. 2 раза > гонадогормонов,
- Мелат. > гонадогормонов,
- Мелат. ³ гонадогормонов,
- Мелат. = гонадогормонов,
- Мелат. £ гонадогормонов,
- Мелат. < гонадогормонов,
- Мелат. < 2 раза гонадогормонов.
Все эти градации превалирования мелатонина или гонадогормонов и определяют становление 7 этапов-ступеней онтогенеза. Очевидно, первые 4 этапа идут под знаком +, когда эпифиз владеет ситуацией, а следующие 3 этапа идут под знаком –, когда ситуация развивается по другим принципам и за пределами оптимума и гормезиса. Очевидно, восьмой этап – это проявление полной девиации эпифиза и гипоталамуса = инволюции. Особенности этапов от 3 до 5, что это переходы от деэскалации эпифиза до элевации гипоталамуса.
Сначала эпифиз сдерживает гипоталамус, затем происходит паритет, а в последующем гипоталамус угнетает эпифиз, причём на каждом последующем онтоэтапе всё больше и больше. Вместе с угасанием эпифиза исчезает этап эпифизарной молодости и максимального уровня Жизненной Силы, нарастает функциональная мощь тканей, количество компенсаторных надстроек и нагрузок на организм.
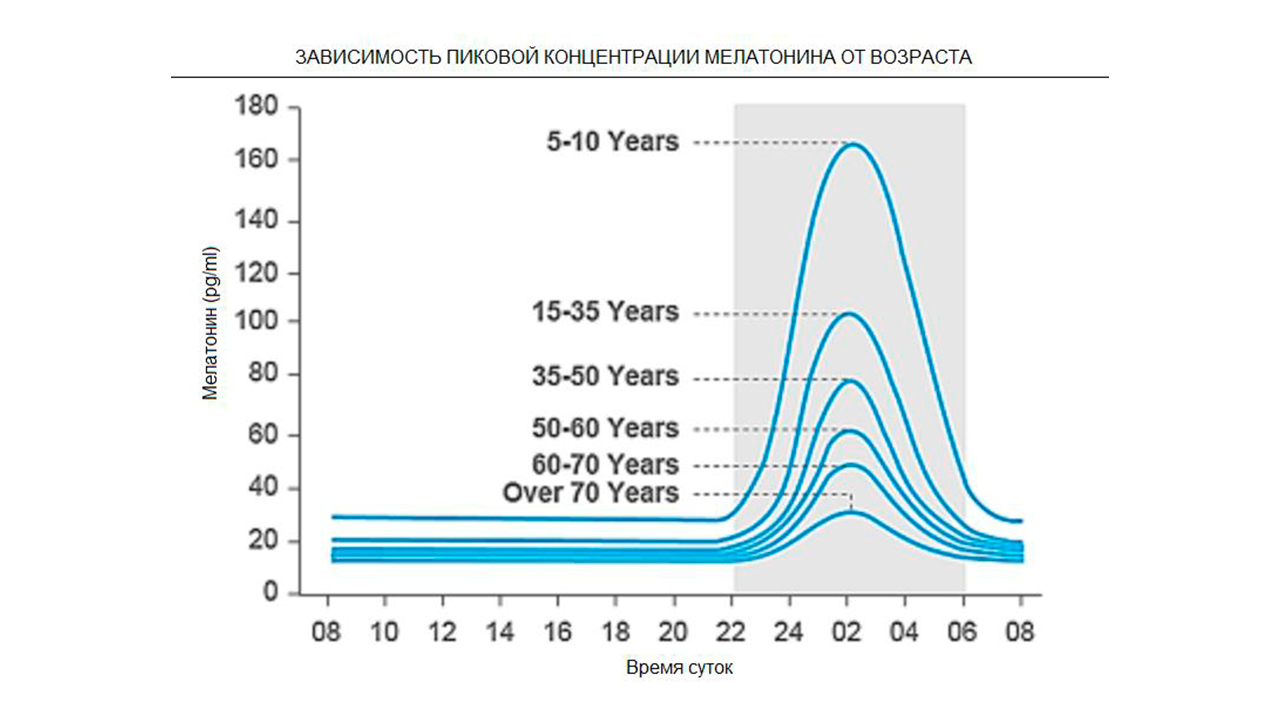
Рис. 42. С возрастом ночной уровень выработки мелатонина неизбежно снижается, что и является драйвером для элевации работы гипоталамуса.
Эпифиз в роли драйвера эскалатора становления этапов онтогенеза, начиная с детства, юности, адальтуса, матуры и сенилита.
Напомню, что этап автодепривации эпифиза – это начало матуры и сенелита, а все остальные этапы онтогенеза являются ступенями становления, которые определяют взаимоотношения эпифиза и гипоталамуса, силы, степени их гормональной соподчинённости, то есть уровня превалирования или паритета того или иного этажа системы горональной регулировки.
Напомню, что выше мы говорили об элеваторной = элевационной функции гипоталамуса, то в случае с эпифизом говорим об эскалаторной функции, где суть этих терминов схожа, но механизмы реализации разные.
Химические формулы мелатонина и цитокинина.
По химической формуле мелатонин состоит из индоламинных веществ, из которых строятся ауксиновые гормоны растений. На этом их сходство заканчивается и действие мелатонина в большей степени проявляется в ювенилизирующем направлении.
Рис. 43. Сравнение формул мелатонина и цитокинина.
Подтверждением этому является феномен эпифизарного доминирования в возрасте до 12 лет, а также возможное участие в реювенилизации у взрослых зрелых особей. Ауксины отвечают за апикальное доминирование. В малых дозах ауксины стимулируют рост корней, а в больших – содействуют зрелости и закладке плодовых побегов. Следует отметить, что многие биологические эффекты связаны с взаимодействием ауксинов и цитокининов. Аналогично, очевидно, и в организме животных, где мелатонин должен взаимодействовать как с некими реювенилизирующими веществами, так и с половыми гормонами.
Высокие уровни мелатонина сдерживают образование гонадолиберинов в гипоталамусе, а значит репродуктивную зрелость. Сдерживая зрелость, они ограничивают степень гипоталамического доминирования.
С этим, очевидно, связано то, что голые землекопы, обитая под землёй в темноте, имеют более высокий уровень мелатонина и более продлённый этап инфантильности = неотении. С этим связано их долгожительство? Если мыши существуют на принципах гипергипоталамического доминирования, то голые землекопы – на принципах эпифизарного доминирования.
Показано, что с возрастом размеры эпифиза увеличиваются двукратно, но связано это с фиброзом, который в свою очередь может быть проявлением длительного провоспалительного процесса. Это означает, что эпифиз находится длительно в состоянии стресса или под чрезмерной сигнализацией обратной связи, которая его стимулирует на первых порах, а затем угнетает. Это аналогично компенсаторному увеличению щитовидной железы при её гипофункции с последующим переходом в гипофункцию?
Отмечено, что это увеличение не связано с уменьшением числа паренхимальных клеток. В пинеалоцитах происходит возрастное уменьшение соотношения объёмов ядра и цитоплазмы, указывающее на гипофункцию клеток. Сделан вывод, что у людей "истинной сенильной атрофии шишковидной железы не происходит". Эпифизы стариков и молодых людей почти не различаются. В клетках эпифиза при старении происходит увеличение числа липофусциновых гранул, инвагинаций ядра, накапливаются кальциевые конкреции, представляющие собой отложения гидроксиапатита, то есть кальциноз. Похоже этот кальциноз связан с возрастной митохондриальной дисфункцией клеток или хронической вегетативной напряженностью. Так депривация симпатического нерва иннервирующего эпифиз животного приводила к тому, что накопления кальциевых конкреций в эпифизе не было. Значит стресс связан с перевозбуждением симпатики.
Таким образом, созревание и усиление вегетативной нервной систем и становление гормональной половой системы деэскалирует эпифиз.
Эпифиз в роли корректора нейроиммуноэндокринной системы.
Очевидно, вместе с тимусом они ведут по ступенькам к старению. Гормон эпифиза мелатонин регулирует пролиферативную активность иммунных клеток тимуса. Верифицированы 4 субпопуляции иммунных клеток эпифиза: недифференцированные лимфоциты (CD5+), Т-хелперы (CD4+), цитотоксические Т-лимфоциты (CD8+) и В-лимфоциты (CD20+). С возрастом число недифференцированных лимфоцитов и В-клеток снижается, тогда как количество Т-хелперов и цитотоксических Т-лимфоцитов достоверно не изменяется. Показано, что эпифиз является органом иммунной системы, а верифицированные в нём иммунные клетки могут выполнять компенсаторную роль при возрастной атрофии тимуса.
Климакс – следствие возрастной и фотопериодической девиации эпифиза.
Если воздействие света на мышей и крыс увеличить до 24 часов в сутки, у большинства из них в короткие сроки разовьется синдром персистирующего (постоянного) эструса. В естественных условиях этот синдром развивается в более позднем возрасте и затем переходит в анэструс, который является физиологическим эквивалентом климакса у женщин.
В яичниках у крыс с персистирующим эструсом обнаруживают фолликулярные кисты и гиперплазию ткани яичника, в них отсутствуют желтые тела. Циклическое производство гормонов, характерное для нормального репродуктивного периода, нарушается, что приводит к гиперпластическим процессам в молочных железах и матке.
Эпифиз верховный корректор чувствительности гипоталамуса, его толерантности, а значит и драйвер онтогенеза.
Деэскалация выработки мелатонина в сторону уменьшения в начальной стадии онтогенеза ведёт к половому созреванию организма, утраты эпифизарной молодости, а последующее снижение его уровня ведёт к увеличению порога чувствительности гипоталамуса, его толенрантности, неподчиняемости, то есть доминантному положению. Гипоталамус становится верховодителем. Он продолжает элевацию, а затем деэлевацию, что проявляется в виде адальтуса, матуры, климакса, а затем сенилита. Но именно деэскалация является причиной деэлевации. Но деэскалация эпифиза является одновременно и следствием элевации гипоталамуса. На первом этапе идёт элевационирование гипоталамуса, а затем – инволюия.
Эпифиз в роли корректора противоопухолевого иммунитета.
Известно, что объём гормона мелатонина, выделяемого эпифизом, играет важнейшую роль в реализации противоопухолевого иммунитета, тем самым играет роль камертона старения иммунной системы.
Экстремальные случаи изменений в эпифизе приводящие к изменению онтогенеза и продления неотении
Случай задержки онтогенетического развития и старения связанный с эпифизом.
Расскажу о Одри Санто, которая вследствие несчастного случая упала в бассейн в возрасте 4 лет. В больницу её доставили в критическом состоянии. Из-за кислородного голодания её мозг получил серьёзнейшие повреждения. Девочка провела в больнице около месяца. Всё это время она находилась в коме. Врачи утверждали, что её жизни ничего не угрожает, но и шансов на выход из комы практически не было. В коматозном состоянии она находилась более 19 лет. Этот случай интересен тем, что в 23 года Одри выглядела как 10-12-летняя девочка. Её рост полностью остановился. Детский невролог утверждал, что работа мозга девочки полностью в норме, несмотря на серьёзные повреждения. Позже врачи изучили тело ребёнка. Оказалось, что она вовсе не в коме. Её состояние оценили как сон. Вот только функции мозга, отвечающие за пробуждение, серьёзно пострадали, и разбудить ребёнка не представлялось возможным.
Учёных заинтересовал феномен, связанный с не взрослением ребёнка. Рост организма, несмотря на получение всех необходимых веществ, серьёзно замедлился. Некоторые исследователи высказывали гипотезу, что в таком состоянии она могла бы жить гораздо дольше, чем обычный человек. Когда ей исполнилось 23 года её мать решилась на исследование головного мозга девочки. Выяснилось, что особенность её мозга представлял эпифиз. Известно, что в эпифизной части мозга у человека к 6-7 годам образуются камнеподобные кальцинаты в области эпифиза, когда клетки его кальцинируются в подростковом возрасте. Причина или назначение этого явления не известны. Ещё одна особенность эпифиза – огромное количество вен и сосудов, которые, словно кокон, покрывают его. По их численности эпифиз стоит на втором месте после почек.
В ходе трепанации черепа и изучения органа выяснилось, что эпифиз девочки не окаменел. Гипоталамус оказался серьёзно повреждён и не выполнял своих функций. Эти два фактора и сказались на не проявлении старения организма. Следовательно, за проявлениями сенилита стоит не гипоталамус, а эпифиз! Теоретически, это означало, что если остановить кальцинирование эпифиза и элевационную гипер-, а затем гипочувствительность (толерантность к сигналам обратной связи) гипоталамуса, зависящую от статуса эпифиза, то физиологическое взросление и, возможно, старение будут отключены. Конечно, можно спорить сможет ли в таком случае человек жить дольше обычного или нет. Но, нейрофизиологи, проводившие исследование, склонны именно к этой точке зрения.
Итак, эпифиз не пострадал, но, очевидно, в связи с тем, что у неё был повреждён центр просыпания и она находилась в непробудном «сне», то были отключены механизмы циркадных ритмов в связи с непрерывным 24-часовым темновым актиноритмом при закрытых глазах, а значит эпифиз не имел преград для непрерывной, персистирующей выработки мелатонина. Уровень мелатонина не колебался ни в зависимости от течения суток, ни в зависимости от сезона, ни в зависимости от возраста и всегда был на предельно высоком уровне. Значит эпифиз стареет не сам по себе, а при факторах подавляющих его. Таковыми могут быть фактор «распакованности» гипоталамуса и медиаторной мощи Вегетативной Нервной Системы, а также таламуса. В принципе он может никогда не стареть, если отбросить из внимания фактор сенесцентности. Именно с этим было связано отсутствие кальцинации эпифиза и выход на сохранение состояния детства с неограниченной пролонгацией неотении*. Переход на очередной этап онтогенеза не произошел. Следовательно, переходы на следующие этапы, ступени онтогенеза связаны со снижением активности эпифиза, то есть его девиацией, подавлением со стороны гиптоталамуса и нейрогуморальной системы, включая вегетативную нервную систему! К сожалению, этот феномен пока только возможен в непрерывном сне. Изученный случай показал, что сенилит не является неизбежной данностью.
«Окаменение» эпифиза, очевидно, и есть тот механизм, который ведёт к его возрастному вырождению, а это в свою очередь разблокирует гипоталамус и ведёт к его зрелости.
Во время этого изучения ствол мозга девочки перестал работать. Врачи диагностировали смерть на операционном столе. Однако, исследование структур головного мозга девочки позволило пересмотреть отношение к эпифизу. Теперь учёные были уверены, виновник старения организма – окаменение отдела мозга.
Некоторые авторы утверждают, что кальций начинает заменять клетки эпифиза в 4-7 лет, а уже в 18-20 лет он полностью превращается в камень. По другим данным в возрасте 5 лет скопления кальция это редкость, которая скорее всего является следствием каких-либо заболеваний. Имеются данные, которые указывают, что активное обызвествление начинается в возрасте около 30 лет. К 65 годам обызвествление обнаруживается почти у 100% людей. Некоторые исследования выявили кальциноз шишковидной железы у младенцев. Это опровергает теорию о том, что старение является фактором, способствующим кальцификации шишковидной железы. Фактически, после этого уже начинается медленное старение организма. К сожалению, современные технологии не позволяют замедлить или остановить этот процесс у человека. Зато ошеломляющие результаты были получены в лабораториях на мышах.
Итак, этот пример показал связь старения с обызвествлением эпифиза. Но, тем не менее, это не объясняет причину и механизм обызвествления! Последовательность событий в этом случае можно трактовать так: первым пострадал из-за гипоксии гипоталамус, в котором находится центр сна и просыпания. Но гипоталамус связан обратной связью с функционированием эпифиза, именно он даёт ему сигнал на созревание, в данном случае этого сигнала не было и эпифиз продолжал функционирование в застывшем режиме без элевации в нём, тем самым сохранился принцип эпифизарного доминирования.
Но не является ли это обызвествление аналогом возрастного отложения белка липофусцина (пигмент старения) в мозговых структурах. Липофусцин — продукт лизосомальной дегенерации митохондрий и в норме постепенно накапливается в цитоплазме клетки в виде гранул, сконцентрированных вокруг ядра. В принципе липофусциноз возможно одно из проявлений сенесцента, но некоторые авторы связывают липофусциноз с ишемией мозга.
Эпифиз - не единственная часть тела, которая может обызвествляться. Кристаллы также могут образовываться в суставах, сердечных клапанах и в ткани молочной железы и на сосудах. Иногда кальцификация ухудшает функционирование пораженного органа. В случае кальцификации шишковидной железы железа может перестать вырабатывать мелатонин. Кальциноз в этих органах можно признать следствием провоспаления.
С помощью специальных химических средств процесс окаменения эпифиза у мышей был значительно замедлен. Жизнь подопытных увеличилась в 6-10 раз! Правда, и взросление, а также половозрелость животных, достигалась гораздо позже обычного срока. Кроме этого, изменениям в структуре подверглись кости скелета. Они оказались более хрупкими, что провоцировало травмы мышей при обычной жизнедеятельности. Наблюдались и сбои в эндокринной системе. Пока научное сообщество делает лишь первые шаги в этой сфере. Возможно, когда-то и правда, благодаря воздействию на эпифиз, удастся увеличить продолжительность жизни человека.
Анализ аномальных случаев не старения человеческого организма в коматозных условиях и последующего резкого старения в короткий временной отрезок.
Описан случай с Иваном Кузьмичом Качалкиным, который под воздействием стресса впал в летаргический сон на двадцать два года.
Случай с Надеждой Артемовной Лебединой в 1954 году. Накануне вечером она крепко поругалась со своим супругом. После чего в раздраженном и расстроенном состоянии отправилась спать. На следующее утро её не смог разбудить уже никто из домашних. Причиной случившегося стал сильнейший психологический стресс. Общим в обоих этих случаях очевидно является то, что глубокий стрессовый шторм пагубно повлиял на центр просыпания в гипоталамусе аналогично тому, что и в случае с девочкой в возрасте 4 лет, у которой произошла гипоксия в гипоталамусе и погиб «центр просыпания».
Примеры с Качалкиным и Лебединой имели ряд общих проявлений, включая то, что тело никак не реагировало на внешние раздражители (при этом кожа бледнела, температура заметно снижалась, пульс почти не ощущался).
В случае с Лебединой, пульс составлял 3 удара в минуту. Внешне человек очень походил на умершего и "дыхание жизни" в нём с трудом улавливалось. Пациент полностью слышал и понимал все происходящее вокруг, но тело ему при этом не подчинялось.
Качалкин «чувствовал страшную, неодолимую тяжесть в мускулах, так что ему было даже трудно дышать». Между тем, он отлично помнил разговоры медсестер в палате. Это был мужчина 60 лет, 22 года лежавший в больнице настоящим живым трупом, без малейшего произвольного движения, без единого слова... Последние годы начал делать движения: в настоящее время встает с постели... говорит много и разумно...».
Аналогично было и с Лебединой. Она также отлично слышала все происходящее вокруг, но все тело было словно налито свинцом и совершенно ей не подчинялось. И как не старалась, она никак не могла сделать хоть какое-то телесное движение. Даже такое простое, как поднять свои веки.
Уточню, что летаргический сон это не кома. Но он же и послужил тем единственным "лекарством", которое вывело пациентов их из него.
Так Лебедина неожиданно очнулась узнав о смерти матери еще в бессознательном состоянии. После "пробуждения" Лебедина прожила еще 20 лет и умерла в 74 летнем возрасте. Качалкин умер в 1918 году в возрасте 60 лет от очередного эмоционального переживания узнав из газет о расстреле царя.
Но важно то, что во время летаргического сна человек не стареет и не меняется как внешне так и внутренне. Время как бы останавливается и тело словно "выпадает" из привычного хода времени и оно становится над таким пациентом не властно. В момент пробуждения такие люди полностью соответствуют тому физиологическому возрасту в котором ранее "погрузились" в состояние летаргического сна.
Как вывод, можно сказать, что время, которое они провели во «сне», не привело ни к каким вторичным последствиям возможных из-за нарушения хода динамики онтогенеза. То есть не сказались возможные проблемы из-за сенесцента, что подтверждает что сенесцент сцеплен с сенилитом. Также не возникли проблемы с неограниченным ростом, который по идее должен был бы продолжаться в фазе затяжного ювенила, ведь ювенил всегда сопровождается открытой фазой роста и гормонам роста, которые сдерживают половые гормоны. Это подтверждает принципиальную возможность реализации продленной ничем неограниченной неотении.
Но есть во всем этом одно весьма существенное "но". Как только человек пробуждается, в очень короткий промежуток физиологическое время его экстренно нагоняет. Человек начинает стремительно стареть до соответствующего его возрасту состояния организма. Период персистирующего непрерывно и в гиперобъёмах функционирования эпифиза мгновенно отключается. Организм возвращается к тому биологическому возрасту, в котором он в этот момент приблизительно должен быть, то есть нет сохранения того периода времени, за который они должны были бы подойти к этой отсроченной онтостадии. Почему так, непонятно. Возможно, всё же шел сенесцент и онтофизис на клеточном уровне при полном отставании процессов сенилита? Но возможно само просыпание является мощным стрессом, который ведёт к преждевременному старению, случаи которого известны в нашей жизни?
Так, Лебедина состарилась менее чем за шесть месяцев. Наблюдавшие её врачи записали об этом: «То, что нам удалось наблюдать незабываемо! Она старела у нас на глазах. Каждый день прибавлял новые морщины, седые волосы».
Во время глубокого летаргического сна организм не стареет. Старение происходит в момент бодрствования (когда человек просыпается).
Во сне в таких пациентах накапливается некая субстанция (сенесцентизм?), которая во время сна ведёт себя пока нейтрально, неактивно. Когда они просыпаются, все её накопившиеся запасы некой субстанции сразу же вступают в работу, начинают активно отравлять и старить человеческий организм.
Но возможно, всё дело в индивидуальных биологических хронологических часах, которые останавливаются на время летаргического сна и запускаются ускоренно после того как организм пришел в состояние бодрствования.
Такой взгляд близок мнению исследователей [1], которые утверждает, что переключение в гипоталамусе происходит на других принципах и выдвигают концепцию, что эта функция переключения на фазу половой зрелости находится в самом гипоталамусе, в котором есть система «Тэнд» – отсчёта времени, а гипоталамус называют гомеостатом времени. Эти авторы предложили ввести понятие о гипоталамусе как регулятора эндогенного времени = Тэнд. Действительно время наступления пубертата и половой зрелости приблизительно удивительно происходит в одном возрасте с небольшими разбросами. Что то жестко «рулит» этим временем. Но это можно легко объяснить ступеньками, рамками в которых идёт непрерывно динамика роста гормонального фона. Имеются жесткие параметры рецепторов в главенствующих структурах как гипоталамус и эпифиз, которые строго следят за этой динамикой и при достижении неких критических точек реагируют включением корректирующих программ. Может это и есть система «Тэнд»? Но правильнее утверждать, что онтогенез это механизм непрерывно движущегося ползунка реостата по реализации неких программ развития. «Ползунком» являются гипоталамус-эпифизарная система.
Тэнд концепция означает, что это система, которая развивается и работает сама в себе, не корректируется общим гормональным фоном. Это означает, что функция регулятора времени изначально должна быть коммитирована генетически, то есть изначально включенной программой для данного типа клеток. Скорее всего это ошибка. Этими авторами не принята во внимание значимость эпифиза в этом процессе!
С позиции концепции «эпифизарного доминирования» можно утверждать, что во всех приведенных случаях произошло не отключение эпифиза. Именно с ним связаны эти «чудеса». Тормозилось обызвествление эпифиза и снятие провоспаления в нём. Эпифиз продолжал доминировать. В гипоталамусе не происходил в достаточной степени рост динамики созревания, а это не позволяло прийти в точку когда гипоталамус стал превалировать над эпифизом и не подавлять его. Но возможной причиной гиперподавленности гипоталамуса является гиперэпифизарное доминирование?
Феномен самоисцеления от рака в коматозном состоянии.
Описаны случаи практически стремительного саморассасывания опухолей даже внушительных размеров. Но особо впечатляет случай самоисцеления пациентки Аниты Мурджани, описанный в её книге-отчёте: «Моя победа над раком». Она заболела лимфомой Ходжкина, (злокачественная гранулёма, лимфогранулематоз, рак лимфоузлов) — это онкогематологическое заболевание, поражающее одну или несколько групп лимфатических узлов, нередко с вовлечением лимфоузлов средостения, брюшной полости и забрюшинного пространства. Характерным признаком его является наличие особых гигантских клеток, обнаруживаемых в поражённых лимфатических узлах. В норме такие клетки должны подчиняться механизмам самообновления и апоптоза, но какой-то фактор, очевидно вирусный, препятствует этому, клетки становятся нечувствительными к сигналинговой гомеостатической саморегулировке.
Болезнь достигла терминальной стадии развития, и завела её в предсмертную глубокую кому и врачи предсказывали считанные дни её жизни. Огромное количество лимфоузлов на шее осложняло её дыхание, ни химия, ни операции не давали никакого эффекта, всё сопровождалось нетерпимыми болями и мучениями с распадом тканей… В предсмертные дни она попала в больницу, где ни один врач не мог предположить её выздоровление. Врач заявил: «У неё уже почти все органы отказали, а по всей лимфатической системе, от основания черепа до живота, опухоли размером с теннисный мяч. Лёгкие и мозг наполнены жидкостью. По всей коже — гнойные раны, сочащиеся токсинами. Она не переживёт эту ночь». Обращаю внимание на факт скопления жидкости в легких, то есть произошло воспалительное выпотевание (экссудативный плеврит) и в мозге, то есть отёк мозга.
Онкология и экссудация жидкости являются проявлением крайней степени мощнейшего воспалительного процесса.
Это означает, что весь её организм был переполнен воспалительными цитокинами (сигнальными молекулами, которые секретируются как иммунными клетками так и самими клетками больной ткани в т.ч. и онкологической), которые могут вызвать локальные некротические процессы или интоксикационный шок ткани с отключением её функции. Воспалительные цитокины преимущественно продуцируются Т-хелперными клетками (Тг) и макрофагами и участвуют в усилении воспалительных реакций. Провоспалительные цитокины (фактор некроза опухоли альфа (TNFα) и интерлейкин 6 (IL6)) играют значительную роль в этиологии и патогенезе как при онкологии, так и при хронике, где индуцируют фиброз. Цитокины осуществляют свои функции в организме через взаимодействие с мембраносвязанными и растворимыми рецепторами. Следовательно, в таких случаях должны быть подключены методы противодействия с избыточностью иммунного и провоспалительного ответов. Это возможно с применением интерлейкинов противовоспалительного направления.
Основной причиной многих онко и хронических заболеваний признан вирус Эпштейна — Барр (ВЭБ).
Он же известен как вирус герпеса человека 4-го типа, который становится своего рода спусковым механизмом патологического процесса.
Вирус Эпштейна — Барр является первым вирусом, связь которого со злокачественными новообразованиями удалось доказать. ВЭБ способен поддерживать латентную инфекцию в организме человека, когда число вирусных копий небольшое и клетка не подвергается разрушению, с периодической реактивацией в течение всей жизни с момента инфицирования. В настоящее время признана ключевая и решающая роль ВЭБ в развитии множества заболеваний, таких как лимфома Ходжкина, лимфома Беркитта, карцинома желудка, носоглотки, агрессивный лейкоз. Ввиду отсутствия у многих таких заболеваний клинических проявлений на ранних их стадиях становится очевидной неэффективность стандартных методов диагностики у большинства пациентов.
В ядре клетки-хозяина ДНК ВЭБ формирует кольцевую структуру — эписому, остающуюся в цитоплазме зараженного В-лимфоцита, или встраивается в геном, вызывая хромосомные нарушения; вследствие этих процессов происходит трансформация В-лимфоцитов в лимфобластные клеточные линии.
В норме цитотоксические Т-лимфоциты и натуральные киллеры (NK-клетки) ограничивают первичную инфекцию и держат пул «бессмертных» ВЭБ-инфицированных В-лимфоцитов под контролем, т. е. экспрессия белков, обладающих трансформирующими свойствами, и белков литического цикла в латентно инфицированных клетках находится под контролем лимфоцитов, активированных ВЭБ.
ВЭБ обладает как оппортунистическими, так и онкогенными свойствами, поэтому он может быть причиной самых различных заболеваний. Найдены доказательства того, что он же является и причиной такого аутоиммунного заболевания как рассеянный склероз.
Все чаще врачи многих специальностей, в т. ч. вирусологи, иммунологи, онкологи, инфекционисты, сталкиваются с развитием ассоциированного с вирусом онкологического процесса без выраженного иммунодефицита. В настоящее время множество исследований проводится по поводу изучения маркеров и многих других показателей для осуществления ранней диагностики предракового процесса, в т. ч. в инфицированном ВЭБ организме.
Возможности мелатонина против вирусов.
Мелатонин оказался эффективным в борьбе с вирусными инфекциями в различных экспериментальных исследованиях на животных и в культуре клеток. Эти данные свидетельствуют о возможном терапевтическом потенциале мелатонина при заболеваниях человека, вызванных вирусом.
Появились доказательства значимости мелатонина в усилении противовирусной борьбы, в том числе гриппа, covid-19, герпес-вирусной группы, к которой относится и Эпштейн-Барра. Следует подчеркнуть важность возможностей мелатонина именно в случаях внутриклеточной вирусной и иной инвазии. Дело в том, что нет лекарств, которые смогли бы подавить их внутри клетки. Но это подвластно мелатонину за счет снятия гликолиза, но запуска апоптоза и аутофагии.
Анализ возможных механизмов самовыздоровления на терминальной стадии онкологии.
Описанная выше терминальная стадия развития в принципе всегда заканчивается мортальным исходом. Но в данном случае процесс пошел по другому сценарию, то есть то, что в быту называют чудом. Через 5 дней метастазы в её теле уменьшились на 70 %. Спустя 5 недель её выписали из больницы. Она полностью избавилась от рака. Очевидно, мозговой отёк, мощная провоспалительная интоксикация и гипоксия мозга, в частности его гипоталамических центров связанных с регулировкой засыпания и просыпания, стали причиной как комы, так и повреждения центров регулировки сна и циркадных ритмов. Развитие комы в данном случае пошло по аналогичному сценарию с другими случаями комы, описанных в данной книге, например, с Одри Санто и др. Особенность случая с Анитой в том, что патологический процесс «зацепил», привёл к депривации некоторых центров и тем самым сместил с нормы работы всю вегетативную нервную систему и ряда центров гипоталамуса, что привело к отмене циркадного ритма работы эпифиза. Он стал работать автономно. Создалась уникальная ситуация сочетания ряда факторов, когда эпифиз перешел на принцип работы в персистирующем (непрерывном, линейном) режиме. Это развернуло динамику процесса из летального направления в противоположное.
В итоге в один из дней длительной комы к ней вернулось сознание, и пошло облегчение. Затем, как по маслу, опухоли начали уменьшаться, улучшилось самочувствие, пошла быстрая положительная динамика, когда в считанные дни опухолевые образования спонтанно исчезли, а все раны зажили.
Это означает, что у организма есть скрытые механизмы автоблокировки онкологии, но они скрыты, заблокированы и задача их вскрыть. Г.А. Гарбузов выдвинул концепт, что такая положительная динамика самоизлечения связана с тем, что открылся «шлагбаум», ограничивающий непрерывную выработку мелатонина. Он пошел потоком, бурной рекой, то есть в супрафизиологических дозах, и за три дня коматоза «смыл» все неверные, заклинившие программы.
В последующем через несколько лет были получены разноречивые научные данные подтверждающие или нет этот концепт.
Так, исследования, где мелатонин применяли в физиологических дозах, не подтвердили предполагаемую его роль как модификатора биологического ответа при сепсисе и гиповолемическом шоке. В одном из исследований задачей было определить, изменяет ли мелатонин высвобождение активированными макрофагами TNF, IL-6 и интермедиатов активного кислорода. Мелатонин в концентрациях от 10(-7) до 10(-4) Мкм не изменяет стимулируемые LPS TNF и IL-6 или стимулируемую PMA продукцию H2O2 и супероксиданиона исследуемыми популяциями макрофагов. Тем не менее, условия методики этого эксперимента не охватили формат условий и состояния, которые возникли в случае с Анитой М.
В тоже время в других экспериментах было показано, что мелатонин вызывает гибель аутофагических клеток при лимфоме Ходжкина, регулируя RORC. Мелатонин в экспериментах проявил противоопухолевую активность при лимфоме Ходжкина посредством ингибирования клеточной пролиферации и стимулирования клеточного апоптоза. Мелатонин увеличивал экспрессию LC3-II и снижало уровень белков p62 при усиленной продукции аутолизосом, что указывает на индуцированную активацию аутофагии. Клетка должна уметь очищаться или избавляться от вирусов. Для этого у неё есть механизмы апоптоза (самоуничтожения) или аутофагии (запуск лизосомальной системы борьбы с вирусами и др. инородными телами, отходными продуктами метаболизма и конформационно поломанными белками).
В последующем появились статьи о механизмах положительного действия и его эффективности при различной онкологии: «Мелатонин в лекарственном лечении больных различными злокачественными опухолями» и др. Результаты очевидны, но скромные. Так показано, что мелатонин может в разы увеличить выживаемость онкобольных, а также повысить эффективность комплексного лечения с 7,5 до 23,5%. По ряду типов заболевания результаты намного выше. Мелатонин рекомендован как дополнение к основному лечению при некоторых типах опухолей, в том числе гормонозависимых. Такие посредственные результаты не удивительны, так как никто не понимал специфику особых условий, когда мелатонин сможет сработать сполна. Экспериментаторы не пытались создать условия персистирования и супрафизиологических доз мелатонина, а также противовоспалительного и антистрессового фона.
Подключение эндорфиновых механизмов как один из компонентов самопомощи организма.
В описанном случае с Анитой, очевидно, содействие оказало и не уходившее особое состояние эмоционального сознания, можно сказать состояние благодати, умиротворенности, эйфории, в котором она находилась будучи в коматозе, которое при этом работало как нейролингвистическое программирование.
Особо следует обратить внимание на то, что в этом особом состоянии у неё самопроизвольно ушли чувство боли, страха, ужаса, немощности. А ведь боль как проявление воспаления провоцирует прогрессию онкологии. Сработали некие механизмы переключения, в том числе и те, которые отвечают за боль, эйфорию, настроение, возможно экстаза? Напомню, что гормонами экстаза являются опиаты в том числе бета-эндорфин. Он выделяется, когда мы ощущаем угрозу или испытываем боль. Тогда он действует как анальгетик. Кроме того, в роли гормона стресса подавляет иммунитет.
Из статьи: «Бета-эндорфины - целостный терапевтический подход к лечению рака» - видно, что бета-эндорфин - это распространенный эндорфин, более мощный, чем морфин, синтезируемый и секретируемый в передней доле гипофиза, он является предшественником проопиомеланокортина. Биосинтез проопикортина в передней доле гипофиза регулируется кортиколиберином гипоталамуса. Очевидно, в описываемом нами случае комы с исчезновением онкоболи произошло расторможение гипофиза с избыточным выбросом бета-эндорфина, что и вызвало состояние эйфории, благоденствия.
Эндорфины обладают различными механизмами действия, такими как обезболивающая активность, противошоковая, противовоспалительная активность, иммуностимулирующая и иммуноподавляющая активность, снимающая стресс активность и вызывающая эйфорию. Главными источниками хорошего настроения являются гормоны счастья - эндорфин и серотонин. Их выработку можно стимулировать наркотиками. Кстати, наркотики (эндорфины) повышают в 5-10 раз выработку дофамина.
Эффект персистирующего режима выработки мелатонина связан с полным дефицитом серотонина?
Серотонин является предшественником синтеза мелатонина и в случае персистирования мелатонина серотонин должен вырабатываться в избыточных количествах. Серотонин снижает болевой порог, обеспечивает полноценный сон.
Но вырабатывается серотонин преимущественно на свету, что, очевидно, было исключено в случае с Анитой, ведь её глаза в коме были закрыты. Для синтеза серотонина солнечный свет абсолютно необходим. Витамин D регулирует переход триптофана в серотонин. Большое его количество действительно прибавит вам радости. Но без серотонина не может вырабатываться мелатонин. Это означает, что в коме серотонина и мелатонина должно быть меньше, так как не запускается мелатониново-серотониновый маятник. Очевидно, замену витамина D осуществил другой механизм? Известен серотониновый синдром, который наступает в результате передозировки приёма антидепрессантов ингибиторного типа, что повышает уровень серотонина. Нет ли здесь связи с некими препаратами, которые ей могли давать?
В целом концепция серотониново-мелатонинового маятника противоречит общему концепту значимости мелатонина в исцелении в описанном случае. Но без мелатонина не могла произойти очистка от вирусов Эпштейна Барр. Следовательно он был.
Возможно, полный дефицит серотонина имеет обратное физиологическое действие, растормаживает обходные, триггерные компенсационные механизмы выработки мелатонина. Но «маятник» при этом не работает, поэтому нет ни циркадных, ни циркадианных ритмов. Идёт только персистирующий режим выработки. Серотонин совместно с мелатонином влияют на циркадные ритмы. Для циркадианных ритмов тоже должны вырабатываться автоматически достаточные их количества. Один стимулирует раскачку выработки другого. Мелатонин синтезируется из серотонина, но без света и витамина D он не вырабатывается, значит не сработает и циркадианный ритм.
Известно, что содержание волонтеров в полной темноте не ведёт к непрерывному сну, а включает внутренний циркадианный режим сна и бодрствования. Значит и в полной непрерывной темноте длящейся неделями и более мелатонин вырабатывается. Это означает, что и в коме возможна выработка мелатонина, причём в завышенных количествах, но без участия механизма в котором задействован серотонин.
Окситоцин – гормон любви – какова его роль?
Определённое подозрение падает также на этот гормон. Из описания состояния больной в коме видно его необычность. Это чувство добра, благодати, умиротворенности… В последующем больная стала сторонницей философии любви. Очевидно и в коме она находилась под влиянием этого гормона. Окситоцин также способен справляться с болью, также он производится во время оргазма и очевидно экстаза.
Какова роль бета-эндорфина?
Выделение эндорфинов способствует повышению уровня гормона роста и уменьшению выработки кортизола (гормона стресса). Способствуют ускоренной регенерации тканей.
Бета-эндорфин - это распространенный эндогенный морфин, используемый для естественного целостного профилактического, терапевтического, стимулирующего и паллиативного лечения рака без побочных эффектов и недорого. Эндорфины играют важную роль в качестве возможных нейромодуляторов в передаче боли, в обезболивании, в толерантности и зависимости, а также в поведении и эндокринных регуляторах, в основном тех, которые связаны с гипоталамо-гипофизарной осью.
Всё это указывает на то, что бета-эндорфин должен обязательно участвовать в комплексном лечении онкологии на фоне одновременного применения мелатонина. Особенно это важно на продвинутых и терминальных этапах онкологии с выраженной симптоматикой боли. В интернет-сетях бета-эндорфин имеется в продаже.
При онкологии на последних этапах особенно важно часто снижать реактивность иммунитета, ввести его в нормальную колею активности. Очевидно, без одновременного применения бета-эндорфина комплексное лечение высокими дозами мелатонина будет на много слабее. При хронических болях выработка этого анальгетика истощается, а значит боль усиливается и постепенно доходит до такой степени, что обезболивающие препараты часто не помогают.
В таких случаях иммунная реакция всегда исковеркана и запредельно гипертрофирована, что ведёт к чрезмерному, неадекватному провоспалению, которое оказывает уже не целебный эффект, а противоположный, стимулирующий болезнь процесс. Известно, что одна из ветвей в медицине развивается в направлении онкоиммунотерапии, но возможности её ограничены и нельзя уповать на запредельные уровни стимулирования иммунитета, палка о двух концах. Иммунка может работать вразнос и только усугубить всё. Идёт самостимулирование заболевания, а это путь к терминальной фазе болезни и мортальному исходу. Как опиат, бета-эндорфин вызывает чувство удовольствия, эйфории. С этим явно связано снижение порога болевого ощущения, боли.
Следовательно, в такой экстремальной ситуации, как например терминальная стадия онкологии, следует в первую очередь снизить уровень реактивности иммунки. Здесь путей влияния очень много, в том числе включая и Куркумн-Адванс, витамин D3 от Витаукт в завышенных дозах. Также в комбинацию лечения следует включить указанный опиат, который также ослабит болевой эффект. Это повысит эффективность мелатонина.
Паранормальные проявления у пациентки в коме.
Также нельзя не обратить внимание на то, что при этом больная как бы «общалась» со своими родными и близкими, слышала голоса врачей помогающих ей. Она приводит реальные факты из жизни близких людей, о которых не могла знать никак. Нечто из паранормальной области. Очевидно, в связи с избытком бета-эндорфина она «получила доступ» к переходу в изменённое состояние расширенного сознания. Возможно, получение доступа стало тем самым «оголением» эпифиза, который стал особо чувствительным к неким информационным каналам? Эти неведомые пока нами каналы связи стали доступными? Явно, на начальных этапах эволюции эпифиз не работал циркадно, а на принципах персистирования. Циркадность это механизм экономичности и более тонкой подстройки к условиям среды.
Так же информацией для размышления могут быть данные об особом статусе людей, которым удалили эпифиз. Они демонстрируют виртуальное би-размещение, при котором они существуют одновременно и в призрачной реальности, и в настоящем. Они существуют в ярком сновидческом состоянии, пока находятся в сознании, и могут чередовать два эти состояния своего сознания. Возможно, организм расценивает это персистирующее состояние эпифиза как его отсутствие? Очевидно, переход с циркадных ритмом на персистирование ведёт к отмене работы аналитического «контролёра реальности» и открывает заслон к виртуальному би-размещению, когда возможно разыгрывать любые психологические сценарии и даже несколько одновременно. «Контролёр реальности» постоянно нас «приземляет» на экзогенные факторы, но не позволяет проходить «эндогенным», которые могут нас объединять с тонким планом. Человек, таким образом, может находиться в двух реальностях одновременно: например, он находится в больнице и идёт в ванную, но по пути он может находиться и в «иллюзорном» мире – на ипподроме, что «останавливает его перед бегущими лошадьми».
Она уверяет что осознала, что причиной её рака была не любовь к себе, да и к другим. Уверена что в коме она осознала свою болезнь и необходимость всеобщей любви.
Видимо в этом состоянии комы полностью отключены гормоны стресса как кортизол, адреналин, норадреналин, ведь они антагонисты мелатонину, но персистируют другие гормоны и нейромедиаторы как серотонин, допамин, ГАМК… Очевидно, на это время произошло перепрофилирование центральных регулировочных систем. Отключение циркадианной работы эпифиза привело к перебалансировке принципов взаимодействия эпифиза и гипоталамуса. В частности они из принципа гипоталамического доминирования перешли на принцип эпифизарного доминирования, а затем на принцип квазиэпифизарного доминирования, то есть персистирующий эпифиз продуцирующий мелатонин в супрафизиологических дозах.
Персистирующее состояние эпифиза – путь к излечению множества хронических заболеваний и сохранению молодости.
Указанный случай является ярким примером полного апоптоза опухолевого процесса, запущенного естественным путём. Что запустило этот процесс в данном случае — не известно, но, очевидно, триггером, спусковым крючком стали процессы связанные с заболеванием и оказавшие тормозящее воздействие на некие центры гипоталамуса. Возможно состояние коматоза было вызвано глубокой воспалительной интоксикацией мозга, когда были отключены центры просыпания (путь к нарколепсии или коме), с которыми связана орексиновая выработка – гормона просыпания, но зато запущены механизмы персистирования эпифиза. Персистирование эпифиза - это уникальное состояние, которое в природе практически отсутствует, но если оно запускается, то делает чудеса, включая от подавления раковых клеток и вплоть до исключения анизомалий*, то есть возраст-зависимых заболеваний, а также сдерживания сенилита. Но важно, что такие механизмы самоисцеления от этой болезни в организме — есть!
Следует отметить, что сочетание выздоровления совпало с периодом в коме. Это навело на ассоциации со случаем Одри Санто, девочки, которая утонула, а спасти удалось не сразу. В результате из-за гипоксии пострадал гипоталамус и в частности центр, который отвечает за просыпание, десятилетия она провела в коме. Но при этом выяснено, что почему-то её эпифиз сохранил ювенильные особенности, не накапливал кальцинаты, а девочка сохраняла признаки детства даже во взрослом состоянии. Произошла своеобразная отмена возможности элевации в гипоталамусе и эскалации в эпифизе, он оставался буквально пожизненно ювенильным, с этим связано и сохранение ювенильности всего тела, так как через гипоталамус была отменена возможность взросления. Возможно, сочетание комы способствовало выработке высокого уровня мелатонина, как в постнатальный период, то есть в 2 и более раз больше чем во взрослом состоянии. В научной литературе, где описываются опыты по применению мелатонина при онкологии, нигде не применялись столь высокие супрафизиологические дозы. Толи чрезмерно высокий уровень мелатонина, толи другие факторы как угнетение центра по выработке орексина (для пробуждения) не давали возможности переключения центра сна на бодрствование; он постоянно был подавлен. Поэтому человек постоянно находился во сне.
Таким образом, науке еще предстоит разработать способы продублирования аналогичного «коматозного» состояния в целях безвредного лечения онкологии.
Попутно подмечу, что определённый уровень стресса, очевидно за счёт выработки кортизола и норадреналина, способны ограничить возможности гиперсекреции мелатонина и могут вывести человека из летаргического сна: именно это и произошло с Качалкиным, когда он в коме узнал о расстреле царя, а также с Лебединой, которая узнала о смерти матери в летаргическом сне. Следовательно, гормоны стресса могут вывести человека из летаргии, но затем это отключит механизмы сдерживания элевации гипоталамуса.
Накапливается клинический опыт эффективного применения мелатонина в комплексной терапии рака различной локализации (груди, простаты, меланомы…) и показан высокий потенциал в лечении. Появились научные данные показывающие повышение эффективности лечения некоторых видов онкологии при сочетании мелатонина с химиотерапией, при этом также повышается выживаемость таких больных, улучшается качество жизни. Признана доказанной зависимость канцерогенеза от нарушений (в том числе возрастных) в работе эпифиза. Можно предполагать, что радикального высокого эффекта в этих экспериментах по применению мелатонина не было достигнуто из-за того, что он не применялся в супрафизиологических дозах и персистирующем режиме, как это было реализовано в случае самоисцеления с А. Мурджани.
Мелатонин содействует выводу онкоклеток из гликолиза.
Гликолиз это особенность бескислородного метаболизма онкоклеток, который они компенсируют чрезмерным употреблением глюкозы. Но возможен такой эффект отмены гликолиза только в больших дозах, а точнее в супрафизиологических дозах мелатонина, чтобы стала возможной программа запуска в онкоклетках на апоптоз. Запуститься такая программа может только в аэробных условиях, то есть на энергетике кислородного типа. Дело в том, что у онкологических клеток практически полностью отключены митохондрии (энергостанции), которые только и могут запустить апоптоз, без митохондрий он не сможет запуститься. Для перезапуска митохондрий нужны специальные стартеры. Кроме того, нужно поменять метаболический субстрат, то есть исключить углеводы, что повернёт метаболизм на путь неоглюкогенеза, что не приемлемо для онкоклеток.
Доказано, что в некоторых патологических клетках транспорт пирувата в митохондрии блокируется. Это изменение метаболизма называется аэробным гликолизом (эффект Варбурга) и часто встречается в солидных опухолях и в других патологических клетках. Переключение с окислительного фосфорилирования митохондрий на аэробный гликолиз даёт больным клеткам преимущества из-за быстрой выработки АТФ и активации пентозофосфатного пути, который обеспечивает поступление нуклеотидов, необходимых для повышенного клеточного метаболизма. Молекулы, называемые гликолитиками, ингибируют аэробный гликолиз и преобразуют клетки в более здоровый фенотип. Гликолитики позволяют осуществлять внутримитохондриальное превращение пирувата в ацетилкоэнзим А. Мелатонин является гликолитическим средством, которое преобразует больные клетки в более здоровый фенотип. Этим можно объяснить, что функция мелатонина как гликолитика объясняет его действия по ингибированию множества заболеваний. Таким образом, общим знаменателем является действие мелатонина по переключению метаболического фенотипа клеток.
Поэтому так важно в онкоклетках отключить гликолиз и запустить аэробизм, такое возможно только на высоком уровне мелатонина и на фоне особых мощных антиоксидантов. Они одновременно являются и веществами гасящими пламя хронического провоспаления, разжигающего гликолиз и стимулирующего рост онкоклеток.
Противодействие гормонам стресса.
Онкология всегда сопровождается высоким уровнем многих гормонов стресса, в т. ч. кортизола, а он подавляет мелатонин! Есть травы и препараты, которые снижают кортизол, например GABA/ГАБА — безопасный природный транквилизатор – есть у нас; а также помогает 1-разовое питание – снижает кортизол. Закажите рекомендации по 1-разовому питанию с целью вызвать аутофагию.
Противодействие провоспалительному статусу.
Онкология всегда сопровождается высоким уровнем многих провоспалительных веществ, которые и разжигают пожар. В частности, к ним относятся некоторые цитокины, которые и вызывают цитокиновый пожар. Некоторые из антиоксидантов, например Куркумин-адванс, но не все обладают и мощным противовоспалительным действием.
Витамин D3 – мощный инфламант, те есть противовоспалительное. Сочетание витамина D3 и Мелатонина могут остановить рост человеческих клеток рака молочной железы, да и др. гормонозависимых опухолей и их метастазирование.
Есть ли у мелатонина максимальная доза и противопоказания?
Многие врачи утверждают отсутствие побочного действия. У любого лекарства существует доза, которая может привести к смерти. У мелатонина такой дозы нет! Также следует взвешивать, что опасней - онкология или текущее заболевание и побочка от неё.
Способы применения Мелатонина.
Мелатонин под названием «МЕЛАКСЕН» или другими можно купить в аптеке.
Для достижения такого эффекта нужны дозы в несколько раз выше тех, что применяются при других назначениях. Доза мелатонина 20–40 мг показала большую эффективность, чем доза 1,5–5 мг. Напомню, что в ночной период, когда мы спим, уровень мелатонина повышается в 10 раз, а это означает, что и искусственное поддержание его высокого уровня практически безвредно. Те отрицательные стороны, которые могут проявится при его применении многократно раз ниже чем последствия от основной онкологической болезни.
С учетом опыта феномена самоисцеления от рака в коме и его персистирующего выделения, автор предлагает принимать его дозировано через каждые 1-2 часа по 1 таблетке 3-5 мг, рассасывая его длительно под языком, что позволит ему быстрее через слизистую рта попасть в кровеносную систему и мозг. Перерыв между приёмами 1 или 2 часа и по 1 или 2 таблетки за приём следует подбирать индивидуально. Перерыв можно допустить утром на 3 часа.
Курсы его приёма проводить по 15 дней, перерыв 10-15 дней и повтор.
Сам гормон мелатонин, очевидно, следует подавать в супрафизиологических дозах и практически постоянно на период лечения или так, чтобы организм находился под его воздействием минимум 20 часов в сутки. Для этого нужны совершенно иные технологии его непрерывной поставки в организм.
Процедуры лучше проводить в кемпинговых условиях, то есть за городом. Желательно при этом поменять режим бодрствования и сна на противоположный, то есть днём спать, а ночью бодрствовать. Но если такой возможности нет, то можно проводить приём мелатонина и в привычном режиме. Подробности в книге автора: «Актиноритмическое лечение…» [13].
Следует принять во внимание, что ночью с 24 до 3 часов уровень мелатонина повышается в 10 раз и особенно это выражено у детей. Это означает, что возможно те те целебные дозы дозы, о которых идёт речь, тоже могут быть заниженными!
Метод кемпинга – как один из способов восстановления работы эпифиза.
Исследователи приводят данные, что после всего лишь одной недели кемпинга без искусственного света (на природе) уровень мелатонина нормализовались на 100%, а всего лишь после одного выходного дня кемпинга без искусственного света, циркадные ритмы нормализовались на 69%.
Более подробно тема корректировки функции эпифиза будет озвучена в последующей рукописи Г. А. Гарбузова: «Актиноритмическое лечение онкологии» [13].
Может ли искусственная (медикаментозная) кома стать аналогом самопроизвольной ведущей к самоисцелению?
Подробности смотрите в статье: «Механизм феномена самоизлечения от рака в коматозном состоянии» на сайте Гарбузова.
Есть ли отрицательная сторона от длительного применения гипердоз мелатонина во взрослом состоянии.
Исследования по кратковременному применению мелатонина (МТ) в больших дозах (до 1,5 г), показало возможность вызова у них сонливости и спазма мышц живота.
Есть мнения, что применение в дозах близких к естественным может увеличить сонливость. Тем не менее, известно, что грызуны, у которых основная активность переходит на ночное время, все же продолжали вырабатывать в ночной период мелатонин и это не мешало их активности.
Известно что выработка МТ сопряжена с выработкой гормона роста – соматропина (СТГ). 90% СТГ вырабатыветя во сне. Оба эти гормона вырабатываются ночью. Очевидно чем больше мелатонина, тем больше гормона роста. Нарушения сна ведут к недостатку гормона роста, поскольку он вырабатывается только в фазе глубокого сна. Но особенность человека в том, что его биология связана с гомойотопией – ограниченностью роста. С возрастом уровень гормона роста значительно падает. Но в нашем случае при увеличении МТ увеличится и СТГ, что может спровоцировать акромегалию, то есть диспропорциональное увеличение роста ряда мягких тканей и хряща, в том числе носа, ушной мочки и др. Рост в высоту и гигантизм при этом не будет происходить, так как закрыты все костные эпифизарные точки роста путём их окостенения. Но практика не подтверждает данные о том, что неотения способствует акромегалии.
СТГ способен омолаживать клетки организма, буквально «возвращая время вспять», помогает восстанавливать дряхлеющие с возрастом мышцы. Так что определённое повышение его уровня полезно.
Кроме того, избыток мелатонина должен тормозить половую функцию. Но примеры животных с продлённой неотенией показывают, что у них сохраняется половая функция и способнсть к размножению.
Гиперэпифизарное доминирование – пролог к неотении и усиления противоопухолевого механизма апоптоза.
Все вышеприведенные случаи были связаны с коматозным состоянием, а также их можно объяснить с состоянием гиперэпифизарного доминирования. Тогда это может означать, что гиперэпифизарное доминирование может стать механизмом для поддержания неотении, то есть пролонгирования состояния детства или юности. Но в последнем случае с Анитой Мурджани произошло и самоисцеление от онкологии. Это тоже можно объяснить с особенностью процессов происходящих в её эпифизе. Очевидно, здесь тоже были запущены механизмы выработки мелатонина в супрафизиологических дозах, то есть многократно раз выше любых физиологических. Кроме того, это привело к уникальному состоянию, когда мелатонин вырабатывался в гипердозах не только ночью, но и днём, круглосуточно, то есть циркадные (суточные) ритмы эпифиза были отменены и он перешел на непрерывное нециклическое функционирование. Это так называемый полный актиноритмический цикл, когда нет светового периода, а есть только темновой. Подробности см. в статье Г.А. Гарбузов «Актиноритмическое лечение онкологии» [13]. Такие гигантские уровни мелатонина (десятикратно) с персистирующим = непрерывным принципом выделения подавляют элевацию гипоталамуса. Гипоталамус всего лишь партитура, на которой разыгрывается симфония. Он реализатор воплощений эпифиза.
Кроме того, такой актиноритмический режим (24 часа темнота и 0 часов свет) выброса мелатонина многократно усилил механизмы апоптоза (самоуничтожения) онкологических клеток. Следует отметить, что именно онкоклетки в первую очередь из-за своего глубокого гликолиза подвержены апоптозу. Апоптозу способствовали огромные токсичные раны, которые запустили механизмы АФК – активные формы кислорода, которые в первую очередь пагубно сказываются на онкоклетках и провоцируют в них апоптоз? Мелатонин же способствовал и усилению выработки NO – оксида азота, у которого свои взаимоотношения с онкоклетками.
Нейромедиаторы антистресса могут корректировать глубину гипердоминирования эпифиза.
К таковым, очевидно, относятся ГАМК – гамма аминомасляная кислота и другие, но в супрафизиологических дозах. В общем коматозный эффект персистирующего эпифиза с гиперсинтезированием можно пробовать достигать искусственно и без самой комы.
Митохондриальный NO сигнальное вещество, модулятор клеточного дыхания.
Он как клеточный гормон помогает исключить аберантные клетки: как онкологические, так и сенесцентные (постаревшие клетки, утратившие способность к делениям). Любые из этих отклонений от нормы имеют в своей основе гликолиз (анаэробный метаболизм), когда функция митохондрий ослаблена. В этом случае клетка входит в репликативный ступор или в другую крайность – репликативную неконтролируемость. В любом случае ликвидирует их за счёт механизмов апоптоза. В ряде случаев его возможностей не хватает, чтобы запустить апоптоз. Здесь ему навстречу может прийти помощь от супрфизиологических доз мелатонина. Оксид азота участвует в регулировке синтеза АТФ, активности митохондриальных АТФ-зависимых калиевых каналов, важный регуляторный компонент активно делящихся клеток, митохондриальной мегапоры и специфического белкового фактора адаптации к гипоксии. В клетках высших эукариот NO — патофизиологический регулятор клеточного цикла, старения и запрограммированной клеточной смерти. О возможностях NO можно познакомиться в книге Г.А. Гарбузова: «Самоуничтожение раковых опухолей. Феномен апоптоза» [12]. Также рассматриваются возможности NO в поддержании молодости клеток в книге Г.А. Гарбузова. «Здоровье, молодость, долголетие» [2].
Уровень оксида азота в организме регулируется образованием мелатонина. Циркадное сопряжение актиноритмических режимов и уровня выработки мелатонина совпадает с суточной динамикой повышения уровня геомагнитной активности, с которой имеется связь со степенью разрушения гранулоцитов крови и с образованием оксида азота, уровень которого снижается.
Следовательно коматозное состояние может привести к разблокированию ограничений функции эпифиза, а затем к супрафизиологической корректировке уровней мелатонина и оксида азота, что в свою очередь может оказать выраженный эффект на проявление неотении, неограниченной юности, а также на выраженный апоптоз онкологических клеток. Таким образом, это может оказаться мощным механизмом подавления как сенилита (общеорганизменного старения), так и борьбы с онкологическими заболеваниями.
Что общего со случаем неотении в связи с гипоталамическим нанизмом и коматозным эпифизарным доминированием?
Случай гипоталамического нанизма (недоразвития гипоталамуса) нами описан на примере Дениса Вашурина. Его фото вы можете посмотреть на рис. 3. Мужчина 32 лет, который выглядит как 14-летний школьник. В данном случае произошло ограничение возможностей элевационирования гипоталамуса на врожденном генетическом уровне. То есть сигналы или рецепторы на дальнейшую элевацию не срабатывают. Это же одновременно не позволяет гипоталамусу подавлять эпифиз. В итоге не подавленный эпифиз проявляет симптоматику эпифизарной молодости или неотении. Хороший неотенический подарок от генетики для долгожительства?! Такая неотения имеет ряд преимуществ перед неотенией коматозного происхождения, которая может быстро закончится при сбрасывании коматозного состояния.
Случай омоложения организма после пересадки печени, то есть показывающий возможность реювенилизации из старости в молодость.
А теперь давайте проанализируем случай омоложения из сенилитного* состояния в адальтус*, то есть это не пролонгирование молодости, а именно возврат. В Индии 61-летний мужчина Кагака Пилла, страдающий от тяжелого заболевания печени, избавился от ряда недугов, а также стал выглядеть моложе на несколько десятков лет, после того, как ему пересадили орган 21-летнего студента.
С 2008 г. умирающий мужчина ждал донора. Врачи пересадили ему печень молодого человека, погибшего в результате несчастного случая. В результате он с годами начал лишь молодеть. Через два года его седые волосы стали обратно черными, а также он избавился от возрастных морщин. Через 3 года больной распрощался с сахарным диабетом, которым он страдал на протяжении 12 лет страдал.
Какой механизм реювенилизации здесь сработал? Все перечисленные выше случаи продлённой молодости были связаны с эпифизом. В данном случае не было смены или отмены онтостадийных переходов. Генеративная половозрелость не изменялась. Очевидно процесс пошел на клеточном уровне путём снятия сенесцента. Молодые клетки печени выделяли секретом с омолаживающим фактором в организм, то есть стали индуцибельным фактором. В данном случае есть некоторое сходство с экспериментом парабиоза (сшивания кровеносной системы молодой и старой мыши), то есть ткани молодого организма индуцировали омоложение тканей старого. Но эффект трансплантации или парабиоза очевидно можно заменить методом биотрона по Цзян Каньжэну или парабиогенеза* по Г.А. Гарбузову, когда в пище преобладают живые корешки от молодых проростков из семян.
Роль эпифиза на конечных этапах онтогенеза и старения
Почему эпифиз стареет первым?
Ткани эпифиза подвержены довольно раннему старению. Причины этого феномена не известны точно, но некоторые ученые полагают, что причина раннего развития гипофункции эпифиза заключается в том, что одну из своих важнейших ролей он выполняет на этапе полового созревания, не позволяя организму расти слишком быстрыми темпами и вступать в фазу зрелости преждевременно. Поле того как эта миссия выполнена, его активная деятельность снижается.
Седые волосы – важнейший маркер старения и какая связь с мелатонином?
Известно, что мелатонин важнейший гормон отвечающий за выработку Меланоцитостимулирующего гормона (МСГ). Это полипептидный гормон, вырабатываемый средней и задней долями гипофиза и отвечающий за выработку пигментов сетчатки глаза и кожных покровов. Секреция меланоцитостимулирующего гормона осуществляется под контролем гипоталамуса и вырабатываемым им гипоталамическим нейрогормоном меланолиберином.
Подача в организм мелатонина седым людям не проявляет прямой эффект восстановления цвета волос. Тем не менее, косвенная связь имеется. Дело в том, что мелатонин является также антиоксидантом и сдерживает оксидативный стресс. Но в луковицах седых волос стволовые клетки меланоциты теряют активность и, очевидно, на их стаус в первую очередь сказывается не сам уровень мелатонина, а уровень воспалительных цитокинов, который с возрастом только увеличивается. Меняется соотношение мелатонин/воспалит. цитокины. Простое повышение мелатонина в крови не устраняет возрастное провоспаление, которое поддерживает и клеточный сенесцент.
Принципы противодействия сенилиту путём поддержания эпифизарной молодости
Метод кемпинга – как один из способов восстановления работы эпифиза
Исследователи приводят данные, что после всего лишь одной недели кемпинга без искусственного света (на природе) уровень мелатонина нормализовались на 100%, а всего лишь после одного выходного дня кемпинга без искусственного света, циркадные ритмы нормализовались на 69%.

Рис. 44. Оптимальным решением было бы проводить такие курсы в условиях кемпинга, на природе подальше от людей.
Более подробно тема корректировки функции эпифиза будет озвучена в последующей неопубликованной книге Г. А. Гарбузова: «Актиноритмическое лечение онкологии» [13].
Магнитотерапия – перспективный метод повышения чувствительности эпифиза и повышения выработки мелатонина
Магнитотерапия повышает мелатонин.
Дело в том, что любая онкология, стресс, старение приводят к уменьшению производства мелатонина, а железа атрофируется. Но сверхважная задача достигнуть устойчивого повышения уровня мелатонина естественным путём. На такую возможность показывают исследования по применению супермощных неодимовых магнитов. Особенность методики состоит в том, что их применять необходимо длительно, чтобы достигнуть лечебного эффекта.
Для этого предложен специальный головной убор, в котором находится до 12 неодимовых супермагнитов. Первоначальная цель исследований шла в направлении подавления опухолей мозга магнитами. Но попутно проявился неожиданно и другой ценнейший эффект по активации выработки мелатонина железой. Это происходит благодаря восстановлению шишковидной железы. Познакомиться можно в статье: «Магнитная выработка мелатонина». Железа якобы содержит мелкие кристаллики магнетита. Этим и объясняют чувствительность железы к магнитному полю. Оно на эпифиз оказывает исцеляющее или скорее даже регенерирующее действие.
Омоложение организма на клеточном уровне.
Особое внимание обращает на себя возможность реального восстановления работы железы эпифиз. Это открытие! И этот метод считаю самым перспективным и естественным методом лечения. Он попросту омолаживает и восстанавливает сломанную или переродившуюся железу. Мелатонин, кстати, является не только гормоном сна, но и гормоном молодости. Он работает в противофазе с серотонином - гормоном счастья и бодрствования. Их оптимальное соотношение и количество особо велико в детстве и юности. Количество мелатонина у стариков в десятки раз меньше, чем у детей. Считаю, что метод магнитотерапии должен быть обязательно подключен к нашей комплексной программе лечения бессонницы. Необходимо знать, что с возрастом выработка мелатонина резко снижается неминуемо, то есть закономерно у всех. Это сопряжено с естественным старением на клеточном уровне, утраты ими функциональной работоспособности, а затем деградации и увядания всех клеток организма, но быстрее всего это проявляется в клетках эпифиза. Это, наверное, единственный способ, который способен омолаживать на клеточном уровне. В норме уровень мелатонина клетки эпифиза корректируют относительно магнитного поля Земли. Это основа первичной регулировки всех клеток. Подробно об этом смотрите в моей книге: «Лечение раковых и дегенеративных заболеваний электромагнитными полями» [17].

Рис. 45. Шапка в каркас которой встроены магниты. Для постоянного ношения при магнитотерапии бессонницы и преодоления старения.
Рекомендуется изготовить деревянный каркас, а по периметру все 12 магнитов. Над лицом открыто. Магниты укрепляют плотно, чтобы не двигались.
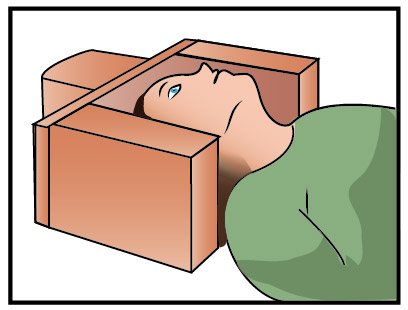
Рис. 46. Изделие из деревянных коробов в которых размещено 12 супермагнитов для лечения бессонницы.

Рис. 47. Внешний вид неодимового магнита.
Для наших целей правильнее будет применить не один магнит, а их целую систему. За основу лучше взять 12 магнитов размером 50 на 40 мм. Сила притягивания такого магнита до 115 кг, а в Гауссах это выражается в 10000 – 12000 Гаусс или 1000 - 1200 мТл.
Северный полюс магнита ориентируют на тело.
Чем отличается клеточное старение (сенесцент) от системного старения (сенилит)?
Общеорганизменное старение или сенилит реализуется через становление этапов развития организма в виде повышения степени зрелости тканей, то есть их онтостадийности, где последним этапом является сенилит. Все этапы реализуются через регулировки в гипоталамусе.
Сенесцент – это другой путь или этаж старения, которое происходит на уровне всех клеток. Он обуславливает лимит Хейфлика, то есть утрату способности их репликации обычно после 52 митозов. У растений такого старения нет, он присущ животным.
Нарушение энергетики клеток или митохондриальная дисфункция как один из элементов клеточного сенесцента.
Обычно он сопряжен с ухудшением работы митохондрий клеток, ослаблением энергетики и частичным переходом на гликолиз. С возрастом содержание кофермента НАД+ уменьшается, из-за чего нарушаются функции нейронной сети и митохондрий (последние обеспечивают клеточное дыхание).
Связь энергетики клетки с электростатическим потенциалом на хроматине и мембранах клетки.
Дело в том, что функцией митохондрий является также поддержание высокого потенциала как на ретикулярной сети клетки, так и в пределах ядра клетки. Особенность высших мета-клеток – создавать и удерживать сверхвысокие потенциалы. Особенно важны эти потенциалы для хроматина высших клеток.
Укорочение теломер является следствием отсутствия реверсного ответа теломеразы на их восстановление по причине её репрессии из-за слабой натяжки хроматина.
Клеточное старение связывают с теломерами, которые при каждом митозе укорачиваются. В принципе они должны бы восстанавливаться, если бы срабатывал механизм фермента теломеразы ответственный за это восстановление. Электрозаряд на хроматине сенесцентных клеток понижен и приводит к изменению электромагнитного поля в ядре: оно резко слабеет. Из-за этого слабеет натяжка на хромосоме, что в итоге приводит к отключению фермента теломеразы.
В онкологических клетках всё очевидно происходит наоборот: из-за избыточного количества активного кислорода АФК митохондрии перезаряжаются чрезмерно, что ведёт к чрезмерному натяжению хромосом. Как итог, происходит эпигенетическое форсирование, экспрессирование гена теломеразы с одновременным включением генов отвечающих за вступление клетки в фазу инициации очередного митоза. Возможно, теломеразная репарация теломера и индукция к митозу – программы сцепленные, зависимые друг от друга, но такая связь возможно только при определённых условиях.
Причина нарушения заряда на хроматине – эпигенетический дисбаланс метилирования на хроматине.
Омоложение через аутофагию
Интервальное питание – путь к идеальному здоровью
Подробности смотрите в одноименной статье Г.А. Гарбузова [9]. Дело в том что интервальное голодание, то есть 1 или 2 разовое питание в день – это путь к притормаживаванию разболтанного гипоталамуса, а значить возврат в рамки коридора гормезиса и восстановления гармонии Витаукта – Жизненной Силы. С возрастом в гипоталамусе снижается чувствительность к сигналам обратной связи и он выдаёт разрушающие нас сигналы, то есть работает за пределами оптимума и гармонии. С этим связано множество возраст-зависимых патологий связанных как с гормональной разрегулировкой, так и нейрогенной. Нарушается гормональный фон, искажается работа вегетативной нервной системы и связанная с нею медиативная регулировка трофики тканей. Усиливаются провоспалительные процессы во всех тканях. За всем этим шлейфом идёт множество возрастных перестроек, по сути вредных для организма и являющихся сутью старения, такие как поседение волос, старческие пигментные пятна, ухудшение зрения, памяти, работоспособности, хроническая усталость, усыхание слизистых, в том числе сухость влагалища, саркопения... Жизненная Сила организма стремительно истощается.
Ключевым словом в данном высказывании является: в гипоталамусе снижается чувствительность к сигналам обратной связи и он выдаёт разрушающие нас сигналы, то есть работает за пределами оптимума и гармонии. Тогда возникает вопрос: откуда возникают начала этой нечувствительности? Ответов на этот вопрос может быть множество, но какой из них правильный?
Для того чтобы правильно разработать технологию противодействия старению, необходимо создать правильную модель его становления. До сих пор научный мир не способен ответить на вопрос откуда идут первичные начала, корни проблемы. Поэтому и существуют десятки мозаичных концепций.
Тем не менее, Гарбузовым Г.А. проведен глубокий анализ природы старения и молодости в растительном мире [3], где чётко показано, что первичные механизмы старения здесь исходят институировано, сверху, то есть на уровне всего организма. У животных – это в принципе тоже ведущий изначальный путь, когда динамика онтогенеза организма выводит его за рамки Великого Гормезиса = сенилиту, который тоже реализуется через механизмы гипоталамуса. Существует еще выход за малый или просто гормезис, который связан с внешними факторами, но не с внутренними механизмами онтогенеза. Тем не менее, на пути у механизмов институирования может стоять множество коммитированных ограничений, которые в свою очередь тоже определяют продолжительность жизни того или иного вида.
Итак, гипоталамус является «поводырём», проводником онтогенеза, через него реализуется продвижение всех этапов организма. Но гипоталамус, в то же время, сам зависим от состояния организма и последнее может сказываться на гипоталамусе, скорости продвижения его выхода за пределы гормезиса. Тем не менее, в этой системе статус гипоталамуса является лидерным, он доминанта! Он коммитативно и институированно определяет темпы старения. Коммитации от внешних тканей и систем, действующих и на гипоталамус, менее значимы. Даже лимит Хейфлика является вторичным и зависимым процессом от той доминанты, которую задаёт гипоталамус.
Казалось бы, при таком ракурсе вмешиваться в такой механизм регулировки сенилита, как вмонтированной фазы развития в онтогенез бесполезно. Раздвинуть онтогенез нельзя, а можно лишь работать в рамках малого гормезиса, то есть бороться с внешними стрессовыми факторами, которые откладывают свой отпечаток на темпы элевации = старения гипоталамуса и могут определять преждевременное старение. Работа в пределах малого гормезиса может реально сдерживать преждевременное старение. Но это влияние не превышает возможности Большого Гормезиса, то есть границ возможностей онтогенеза, где последним его этапом реализации является сенилит. Можно ли обойти рамки Большого Гормезиса остаётся открытым, спорным вопросом. Попытаемся разобраться в этом вопросе и ответить на него оптимистично. Но сразу предвещу, что сделать это можно только через взлом программ и запуск механизмов реювенилизации. В растительном мире эти методы реювенилизации более менее понятны и доказывают их реальную возможность на неограниченное существование. Как осуществить полную реювенилизацию в мире животных и возможна ли она в принципе? На этот вопрос попробую ответить и даже положительно.
Опыт профессора Йошинори Нагумо по омоложению организма на 20 лет с помощью 1-разового питания
Убедительным доказательством в правильности его подхода в стимулировании аутофагии и за счет этого глубокого омоложения организма является его личный опыт, который в 60 лет выглядит на 40-летнего.
Он своей внешностью на фото убедительно разбивает миф о целесообразности трёхразового питания.

Рис. 48. Профессор Йошинори Нагумо в возрасте 60 лет, у которого отсутствует симптоматика старения.
Эту диету он применяет уже 20 лет. Врачи-диетологи ортодоксы не могут поверить и объяснить его феномен. Это противоречит всем канонам сложившейся диетологической науки. Поэтому об этом стараются умолчать или забалтывают различными рассуждениями.

Рис 49. Внешность профессора в 59 лет выглядит на много моложе чем в 36.
Профессор утверждает, что этот метод является эликсиром вечной молодости.
Его методика построена на полном исключении чистых углеводов и сахаров. Фрукты и ягоды должны быть ограниченными и лучше в цельном виде с кожурой. Избегать омега-6, трансжиры. Преимущество льняному семени. Избегать всяких солений и промышленных приправ, чайные, кофейные и другие напитки. Сторонник ферментированных продуктов.
Конечно, здесь рано еще обсуждать на каком уровне аутофагия смогла сработать: на уровне Малого Гормезиса или Большого Гормезиса. То есть здесь удалось устранить преждевременное старение или раздвинуть онтогенез? По крайней мере, ясно одно, что аутофагия будет отличным подспорьем методам медиации по типу цитокино-цитокининовым препаратам с целью восстановить полноценно точки самообновления = точечной реювенилизации.
Также вы можете познакомиться с методами омоложения организма через корректировку сна в статье Г.А. Гарбуозов: Омоложение во сне: проснись ребенком [16].
Термины:
Адальтус — этап онтогенеза после юности, то есть генеративной зрелости.
Адаптационный Универсальный Клеточный Ответ (АУКО) — факторов клеточного стресса бывает множество, но механизмы их стресса и итоговые последствия тоже универсальны, сходны. Автором предложено обозначить как как АУКО по аналогии с стрессом или Общим Адаптационным Синдромом (ОАС). Понятие АУКО в биологии ввел впервые Гарбузов Г.А.
Анизомалия — неадекватные для Витаукта параметры жизнедеятельности, которые находятся за пределами коридора гормезиса, которые ведут к неизбежным особым состояниям и которые накапливаются с возрастом. Понятие анизомалия следует обязательно отделять от понятия возрастные болезни, так как при болезнях обязательно надо уделять внимание на конкретные механизмы патологии и локальное воздействие на них, что абсолютно не подходит при анизомалиях, то есть возрастных перестройках связанных как с сенесцентом, так и сенилитом всего организма, при которых в первую очередь нужны совершенно другие методы, в частности общего омоложения и оздоровления организма (повышения Жизненной Силы противодействующей сенесценту) и укрепления его Витаукт (противодействующего сенилиту), иначе все остальные частные методики будут не эффективны. Аналог устаревшему термину возрастзависимые «болезни». Анизо — перекос, несоответствие, «малия» — часть от слова аномалия, то есть отклонение от нормы; термин предложил Г.А. Гарбузов.
Витаукт — дословно Жизненная Сила, сила или программы, сохраняющие интегрированный оптимум всех гомеостазов, поддерживающая идеальное здоровье.
Витахора — следует понимать, как Жизненная Сила, источник сил которой находится за пределами материальных механизмов организма, его коридора гормезиса, но которые могут влиять на механизмы регуляции Витаукта. Термин предложил Г.А. Гарбузов.
Геронтос — это единство сенесцент ⇆ сенилит. Общее старение которое исходит как со стороны сенилита, так и со стороны сенесцента.
Гипоталамическое доминирование — когда динамика онтогенеза полностью подчинена регулировке гипоталамуса. Такое состояние наступает после прохождения онтоэтапа детства и юности.
Гироскоп онтогенеза — в авиации это прибор, способный указывать и реагировать корректировкой на отклонение от заданных параметров курса; но в нашем случае имеется в виду механизм, который ориентирует организм о ходе динамики онтогенеза и при достижении определённых параметров отклонения включающий корректирующий сигнал на возрастные перестройки в гипоталамусе, то есть переход на следующую онтостадию. При этом в случае разблокировки гипоталамуса происходит обратное подавление работы эпифиза; термин предложил Г.А. Гарбузов.
Гомойотопия — ограниченность роста (от слова «гомойо» — способность всех особей вида или органов в них достигать жёстко заданных величин, например размеров, а «топо» — от слова «топология» — размеры, пространство). Термин предложил Г. Гарбузов.
Гормезис — интегрированный свод гомеостазов, задающий коридор оптимума для проявления жизнедеятельности в живой системе, который имеет верхние и нижние параметры регулировки, регулируемые различными противоположными генными программами; центральные параметры регулируют транспозиты, а крайние пограничные — контрпозиты. Термины предложены Г. Гарбузовым. Следует выделять понятия Большой Гормезис — интегральный, свод механизмов, которые определяют предел, границы возможностей динамики онтогенеза организма, в том числе при выходе его за рамки Великого Гормезиса, ведущего к сенилиту, который тоже реализуется через механизмы гипоталамуса; а также малый гормезис — = просто гормезис, то есть интегральная возможность противостоять всем внешним стрессовым факторам, которые откладывают свой отпечаток на темпы старения гипоталамуса. Но это влияние не превышает возможности Большого Гормезиса. Понятия ввёл Г.А. Гарбузов.
Гомеорез — состоит из слов «гомео» — постоянство и «рез» — смещение, сдвиг. Состояние, когда устойчивый гомеостаз может перейти в системе на новый уровень стабильности, констант.
Гормезиорез — состояние, когда параметры устойчивого гормезиса претерпевают возрастные изменения. Термин предложил Г. Гарбузов.
Институирование — особенность закрепления морфологических признаков в растительном мире, регулируемая через изменение гормонального статуса в их апексах или камбиальной ткани, в отличие от животного мира, где главной их особенностью является коммитирование их признаков — биологический смысл этого термина ввёл Г.А. Гарбузов.
Квазиэпифизарное доминирование – это состояние персистирующего принципа функционирования эпифиза или с гиперпродукцией мелатонина не менее двухкратно выше нормы в среднем возрасте; при этом полностью исключена возможность паритета или переход на гипоталамический принцип доминирования. Очевидно, это один из вариантов возможных механизмов сохранения неотении — биологический смысл термина ввёл Г.А. Гарбузов.
Контрпозиты — комплекс программ и генов, направленных на сужение в системе защиты, ограничение их возможностей, срабатывающих в ситуациях зашкаливания.
Коммитированные клетки — то есть с обязательством, внутренним «предписанием» производить только один тип клеток, включены только на одну потенцию, например, унипотентные клетки или выполняют только один функционал или морфогенную единицу.
Матура — этап онтогенеза, перезрелости, климакса с выходом за пределы, границы оптимума коридора гормезиса, предшествующий сенилиту — биологический смысл этого термина ввёл Г.А. Гарбузов.
Матрикальный — главный верхушечный побег, например, у хвойных деревьев, создающий прямоствольность, ось ствола. Термин предложил Г. А. Гарбузов.
Методы деэпигенизации – методы которые снимают эпигенетическую нагрузку приобретаемую клетками при старении и онтофизисе.
Неотения — способность организма продлевать свои ювенильные этапы существования и тем самым продлевать жизнь.
Нормоконстанта – каждый гомеостаз клетки состоит из множества констант, то есть параметров, которые определяют оптимальную жизнедеятельность клетки.
Онтодианность, онтодиана, онтодианная линия жизни – показатели длительности онтогенеза, на которую расчитан данный вид, исчисляемый в годах. Как в растительном, так и животном мире имеются виды, где одни из них проявляют короткожительство, а другие – долгожительство, которое обеспечено у каждого конкретного вида своим механизмом регулировки онтогенеза. Это означает, что у каждого вида длительность жизни изначально коммитирована, то есть генетически запрограммирована. Термин созадан по аналогии с циркадианность, когда ритмы жизни связаны не с внешними факторами, а внутренними. Термин предложил Г. А. Гарбузов.
Онтостадия, онтостадийность – от слов онтогенез и стадии, то есть возрастные ступени по которым проходит онтогенез.
Онтофизис – подразумевает у растений не простое закрепление у клонов неких особенностей роста и плодоношения, а проявление более общего процесса, то есть сохранения степени онтостадийной зрелости, включая высшую степень онтостадийности, когда полностью заблокирован рост или плодоношение на нижних побегах. В отличие от высшей степени онтофизиса у побегов растений в животном мире аналогом этому является сенесцент у клеток. Онтофизис = сенесценту. Разблокировать такое состояние у растений возможно только при особых условиях реювенилизации, то есть снятии апикального доминирования и обязательном превалировании цитокининового фона. При этом расширено понимание этого явления не только для клонов, но и в пределах данной особи растения, когда новое поколение побегов в пределах одного и того же дерева все проявляют продвинутую физиологическую или морфологическую стадийную зрелость. Программы на ювенильность у них заблокированы. Если расширять понимание этого термина на животных, то подразумевается закрепление неких возрастных и онтогенетических перестроек, которые сохраняются и у линий клеток в культуре ткани, аналогично тому, как это сохранение происходит и у клоновых потомств в виде топофизиса (особенностей роста: ортотропного или плагиотропного) или циклофизиса (способность к закладке или не закладке тех или иных генеративных органов), или онтофизиса (с сохранением темпов клеточного роста или самообновления или их полной блокировки). Особенностью клеток животного типа является то, что до сих пор не найден гормон реювенилизации сенесцентных или онтофизисных клеток, что и является причиной «непреодолимости» лимита Хейфлика. При этом особенностью выращивания клеток в культуре ткани является быстрый «перескок» через все онтостадии и остановке на крайней онтофизисной стадии. Термин предложил Гарбузов Г. А.
Феноптоз – тоже что и клеточный апоптоз, то есть запрограммированная смерть клеток, только на уровне всего организма.
Парабиогенез — от слов биогенез, которое в эмпирическом обобщении понимает, когда одно живое активирует, оживляет другое, а слово пара – (дословно рядом, около) означает что активация происходит за счёт живого начала, но не близко видового происхождения – термин предложил Г. А. Гарбузов.
Парабиоз или гетерохронический парабиоз — когда сшивают кровеносные системы молодой особи мыши со старой, при этом происходит омоложение старой и увеличение продолжительности жизни, но состаривание молодой.
Пойкилотопия — неограниченный тип роста (от слова «пойкило» — различный, меняющийся в зависимости от внешних факторов; «топо» — пространство, размеры, которые занимает организм). Термин предложил Г. Гарбузов.
Прогениторные клетки — стволовые клетки, узкоспециализированные только для дифференциации одного типа клеток, это концевая = терминальная ветка потентности для данной ткани и развития после трансформации из плюрипотентных, проходя при этом череду разных степеней потентности.
Пубертат — онтостадийный этап перехода от детства к генеративной зрелости в молодости и адальтусу.
Радиксальное доминирование — от слова радикс – корень, этап онтогенеза когда доминирует гормон корней цитокинин, который и определяет особенности этого онтогенетического этапа. Термин предложил Г. Гарбузов.
Радиксальный тип онтогенеза — тип онтогенеза, когда доминирующим и определяющим является корневой фактор – гормон цитокинин. Термин предложил Г. Гарбузов.
Реостат хроноса — термин предложен Г.А. Гарбузовым как альтернатива термину эпигенетические часы, которых как таковых нет, а есть механизм продвижения по некому реостату реализации определенных программ и механизмов, эта реализация идёт с определённой последовательностью и скоростью. Это более правильно определить как реостат последовательной хронологической реализации онтофизисных фенотипов и паттернов на онтодианной линии, то есть последовательности возможных сценариев развития.
Рецессия клеточная — спад функциональной активности клетки.
Сенесценс = сенесцент, сенесценсные клетки — репликативное старение, которое зарождается на уровне клеток.
Сенилит — старение, которое зарождается на верхнем общеорганизменном уровне.
Стем-доминирование – это когда преобладает стимуляция стволовыми клетками общего развития организма, то есть с низу, а не с верху. На последних этапах онтогенеза матура, адальтус и сенилит, то есть в период инволюции гипоталамуса, его определяет принцип стем-соматического или прогениторного доминирования, когда исчерпаны возможности высокопотентных стволовых клеток, а доминируют соматические и концевые типы стволовых (прогениторные), с минимальной степенью потентности к репарациям, которые не позволяют дальнейшему самовосстановлению его. Понятие стем-доминирование в биологии ввёл впервые Гарбузов Г.А. Stem = стволовой.
Хейфлика лимит — связан с репликативным старением, сенесцентом клеток, которые в культуре клеток предопределены на ограниченное количество делений.
Транспозиты — комплекс программ и генов, направленных на противодействие, расширение, усиление возможностей системы самозащиты клеток.
Циркадный – суточный ритм сна и бодрствования регулируемый внешними факторами.
Циркадианный – ритм сна и бодрствования регулируемый изнутри, изначально заданный.
Эпифизарная девиация — период эпифизарного вечера, заката (отклонение от оптимального уровня активности, старости), который наступает после эпифизарного утра, эпифизарной молодости. Термин предложил Г. Гарбузов.
Эпифизарное доминирование — состояние молодости или когда этот этап онтогенеза, то есть молодости и детства находятся в первую очередь под контролем эпифиза, а не гипоталамуса. Термин предложил Г. Гарбузов.
Эпифизарная молодость — этап онтгенеза регулируется на принципах эпифизарного доминирования. Термин предложил Г. Гарбузов.
Эпифизоскоп, эпифизоскопия — это функция эпифиза реагировать и запускать коррективы на отклонение гормонального фона.
Список литературы
- М. П. Чернышева, А. Д. Ноздрачев. Нейроэндокринный гипоталамус как гомеостат эндогенного времени.
- Г.А. Гарбузов. Здоровье, молодость, долголетие. 2023 г.
- Г.А. Гарбузов. Биология Молодости. 2020 г.
- Г.А. Гарбузов. Неотения – продлённая молодость, можно ли её сохранять неограниченно с помощью аутофагии?
- Г.А. Гарбузов. «Гипосенсорная камера (камера флоатинга)»
- Г.А. Гарбузова. Яйца – овотерапия, диета, которая не просто сдерживает старение, а омолаживает.
- П. М. Маслюков, А. Д. Ноздрачев. Гипоталамические механизмы регуляции старения. 2021 г.
- В.М. Дильман. Большие биологические часы.
- Г.А. Гарбузов. Омоложение через аутофагию. Интервальное питание – путь к идеальному здоровью.
- Г.А. Гарбузов. Ювенология. Гипоталамус – верховный драйвер онтогенеза: Ключ к молодости и старению.
- В. Х. Хавинсон, А. Г. Голубев. Старение эпифиза. "Успехи геронтологии", 2002г., выпуск 9
- Г.А. Гарбузова: «Самоуничтожение раковых опухолей. Феномен апоптоза» 2023.
- Г.А. Гарбузов «Актиноритмическое лечение онкологии».
- Г.А. Гарбузов. Витахора — нематериальная потенция горметической модуляции Жизненной Силы. 2023
- Г.А. Гарбузов. Имажинация – метод самопрограммирования здоровья.
- Г.А. Гарбузов: Омоложение во сне: проснись ребенком.
- Г.А. Гарбузов. Лечение раковых и дегенеративных заболеваний электромагнитными полями.
 Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента, идущих с уровня клеток, и процессов сенилита, идущих на системном уровне. Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенилита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые, по сути, не являются болезнями, а естественными анизомалиями, то есть механизмами реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), о чем автор расскажет в своей следующей книге: «Биология молодости».
Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента, идущих с уровня клеток, и процессов сенилита, идущих на системном уровне. Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенилита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые, по сути, не являются болезнями, а естественными анизомалиями, то есть механизмами реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), о чем автор расскажет в своей следующей книге: «Биология молодости».
Механизм проявления неотении и возможные методики её реализации будут рассмотрены в последующей статье:
«Природа и суть неотении; методы её реализации с помощью аутофагического омоложения».
 Автор впервые показывает, что онкологию можно и нужно победить не нанося организму вред. Оказывается для этого есть скрытые рычаги управления. Таковыми являются программы на апоптоз – самоуничтожение больных и ненужных организму клеток. Особенность онкоклеток в том, что в них эти механизмы не поддаются самозапуску. Тем не менее, программы нужные для этого имеются, но «законсервированы». Задача их вскрыть. Реализовать это можно и нужно через многократное усиление и сочетание программ и сигналинговых путей клеток на оксидативный и нитрозативный стрессы. Оба эти пути являются клеточными медиаторами апоптоза запускающими ферментный каскад, который изнутри как «ножницы» уничтожает больные клетки. Известные на сегодняшний день методики были односторонними, так как затрагивали только сторону оксидативного стресса. Последний может подавлять опухоли, но не даёт радикального результата. Опухолевые клетки имеют степень свободы маневрировать и приспосабливаться, тем самым легко глушить апоптоз. Отрезать «путь к отступлению» возможно только путём запуска противоположно направленного механизма нитрозативного стресса. Только при их взаимодействии возможен запуск полномасштабного феномена апоптоза и капитуляции опухоли. Дополнительно предложен метод как «разоружить» опухоль от самозащиты. Для этого предложена двухфазная тактика подавления опухоли, когда в Фазу 1 группируются методы для перекрытия механизмов агрессивности и самозащиты опухолей, а на Фазе 2 создаются экстремальные условия избирательные для онкоклеток, когда они способны запустить механизмы самоуничтожения. С учетом этих принципов разработана Комплексная Программа.
Автор впервые показывает, что онкологию можно и нужно победить не нанося организму вред. Оказывается для этого есть скрытые рычаги управления. Таковыми являются программы на апоптоз – самоуничтожение больных и ненужных организму клеток. Особенность онкоклеток в том, что в них эти механизмы не поддаются самозапуску. Тем не менее, программы нужные для этого имеются, но «законсервированы». Задача их вскрыть. Реализовать это можно и нужно через многократное усиление и сочетание программ и сигналинговых путей клеток на оксидативный и нитрозативный стрессы. Оба эти пути являются клеточными медиаторами апоптоза запускающими ферментный каскад, который изнутри как «ножницы» уничтожает больные клетки. Известные на сегодняшний день методики были односторонними, так как затрагивали только сторону оксидативного стресса. Последний может подавлять опухоли, но не даёт радикального результата. Опухолевые клетки имеют степень свободы маневрировать и приспосабливаться, тем самым легко глушить апоптоз. Отрезать «путь к отступлению» возможно только путём запуска противоположно направленного механизма нитрозативного стресса. Только при их взаимодействии возможен запуск полномасштабного феномена апоптоза и капитуляции опухоли. Дополнительно предложен метод как «разоружить» опухоль от самозащиты. Для этого предложена двухфазная тактика подавления опухоли, когда в Фазу 1 группируются методы для перекрытия механизмов агрессивности и самозащиты опухолей, а на Фазе 2 создаются экстремальные условия избирательные для онкоклеток, когда они способны запустить механизмы самоуничтожения. С учетом этих принципов разработана Комплексная Программа.
Вы можете её заказать
 Неопубликованная книга Гарбузова Г.А.: «БИОЛОГИЯ МОЛОДОСТИ», смотрите на сайте: garbuzov.org.
Неопубликованная книга Гарбузова Г.А.: «БИОЛОГИЯ МОЛОДОСТИ», смотрите на сайте: garbuzov.org.
Все мы хотим жить на много больше чем имеем возможность, но при этом в молодости, а не большую часть жизни в болезнях и немощной старости. Какова природа стадий онтогенеза и в частности молодости и можно ли её продлевать за счёт укорочения старости? Действительно природа показывает возможности неотении, то есть растянутой молодости. Может ли человек воспользоваться этими же принципами и механизмами? Оказывается да, но здесь есть глубокие законы природы, которые надо понимать и управлять ими.
Среди важнейших понятий определяющих онтогенез автор впервые показал, что он зиждется на принципах онтофизиса и онтостадийности, которые и являются движущими силами. Сделан глубокий экскурс во всю биологию развития живых организмов, что бы «раскопать» принципы, по которым можно строить методики сохранения молодости.
Важным на этом пути, чтобы получить ответы на главные вопросы, необходимо было ответить откуда идут первичные истинные начала переходов на новые стадии и старение. Существовала путаница с каких этажей организма исходят истоки старения и почему стадия молодости столь коротка. Чтобы ответить на эти вопросы пришлось создать концепцию матричных осей, на которых происходит реализация развития организма.
Показано, что существует два направления развития старения: нижний – сенесцент на уровне клеток и тканей и верхний - на уровне общеорганизменных нейрогормональных регулировок – сенилит, которые имеют разные механизмы, но являются двумя рычагами единой матричной системой. Эти потоки сливаются в единый – геронтос, которые проявляется внешне как симптомокомплекс болезней старости.
На основе этих теоретических посылов автором предложены новые принципы запуска матриц развития на неотению, то есть продления молодости, а значит и на преодоление болезней старости.
 Автором дано новое обоснование проблемы онкологии. Это может быть переломным моментом для пересмотра всей устоявшейся теоретической базы канцерогенеза сложившейся в медицине. Предложена новая концепция, позволяющая не только объяснить, уложить в единую картину все противоречивые факты, пазлы, но и предложить универсальную методику лечения, основанную на естественных принципах самоизлечения.
Автором дано новое обоснование проблемы онкологии. Это может быть переломным моментом для пересмотра всей устоявшейся теоретической базы канцерогенеза сложившейся в медицине. Предложена новая концепция, позволяющая не только объяснить, уложить в единую картину все противоречивые факты, пазлы, но и предложить универсальную методику лечения, основанную на естественных принципах самоизлечения.
В своей концепции автор учел, что онкология всегда сопряжена с саморазгорающимися провоспалительными процессами, что в конце концов и является причиной в большинстве случаев мортального исхода при онкологии. Механизмы провоспалительных процессов завязаны с механизмами десинхроноза. Десинхроноз, то есть механизмы высшей разрегулировки и дезинтеграции всех процессов, и меры, препятствующие его проявлению, могут существенно корректировать онкологические процессы в худшую или лучшую сторону. Выяснилось, что глубокие погружения в состояния сна и даже комы могут разблокировать онкологические программы на уровне клеток. Анализ этих случаев и природы их проявления позволило автору утверждать, что онкология по своей сути подобна «зависшим программам» в компьютерах, то есть блокировке. Дело в том, что при онкологии в ряде случаев создаются ситуации на уровне клеток, когда они переходят на энергетику глубокого гликолиза, а в тоже время на их мембранах происходит операторный сбой – не включаются или сломаны регулировочные рецепторы, а точнее сломан сенсорный дисплей, отвечающий за тип энергетики клетки. Такая клетка может войти в тупиковую ситуацию, из которой она не может выйти. Для этого ей нужно запустить специальные репаративные программы, позволяющие ей вновь запустить отключенные высшие программы дифференциального уровня.
Казалось бы, полный тупик – на уровне клетки она не может сама за счет внутренних программ вывести себя из этого состояния… Такие программы клеткой не предусмотрены. В компьютерах это все заканчивается «зависанием». Причин зависания от ошибок много, и мы не можем их все предусмотреть. В этом случае нам остается единственный путь — это стереть все предшествующие оперативные процессы, в которых запрятана ошибка. В пределах организма в онкологизирующих его клетках, где зависли программы дифференциации клеток и сбой их типа энергетики, тоже можно «стереть», смыть эти операционные ошибки и генетические «поломки». Оказывается, такой возможностью обладает ночной гормон сна – мелатонин. Но возможность такого механизма «стирания» может сработать только при определенных условиях – это персистирующая (непрерывная) выработка мелатонина на фоне подавления активности серотонинового крыла серотониново-мелатонинового маятника, и таким образом устранения самовозбуждающихся провоспалительных процессов, пожара реализуемого самими же онкоклетками. Одно крыло этого маятника возбуждает онкопроцесс, а другое – его гасит. Невозможно работать только на одном крыле маятника: одно должно быть запущено на полную мощь, а другое полностью выключено. «Мутации» в том понимании, которое их трактует современная наука, всего лишь следствие, вторичные процессы поломок, по сути это в большинстве случаев аберрации. В норме организм справляется с ними с помощью репаративных механизмов.
 В книге представлена уникальная методика лечения онкологии и ряда неизлечимых заболеваний, в исходе которых происходят дегенеративные процессы. Особенностью этого метода является то, что он зарождался не в лабораториях научных институтов, а исключительно сообществом специалистов энтузиастов и добровольцев больных, которые потеряли надежды лечения в официальной медицине и испытали этот метод на себе. Большинство из них добились такого положительного эффекта, которого невозможно достичь всеми современными методами. Особенность метода заключается в том, что он действует не на химическом или физическом уровне клеток, а действует адресно на истинные механизмы перерождения на клеточном уровне – основной причины множества хронических и онкологических заболеваний. Оказывается первичные причины перерождений клеток связаны с нарушениями механизмов регулировки клеток, которые находятся не в ядре клетке, а на многочисленных сенсорных структурах клетки, так называемых интегральных белках ответственных за целостность электрического контура. Глобальная, главная ориентировка регулирования у них происходит не химическим путем, а исключительно на голограммное поле клетки, так называемый электромагнитный каркас клетки. Это так называемое генетическое поле присуще только высокодифференцированным клеткам. У онкологических клеток это поле отсутствует и происходит дезинтеграция всех внутренних структур клетки. Автором книги впервые дана новая концепция электромагнитной саморегулировки, самонастройки клетки как сложной многоэтажной системы. На основе этого построено объяснение действия имплозионного поля прибора на повышение уровня интеграции работы всех структур клетки. Аналогов этому нет. Дано объяснение механизма репарации больных клеток и исчезновения опухолевых. Проанализирован опыт излечения самых различных ранее неизлечимых заболеваний и дана новая комплексная методика их лечения новыми методами, которых нет еще в медицине.
В книге представлена уникальная методика лечения онкологии и ряда неизлечимых заболеваний, в исходе которых происходят дегенеративные процессы. Особенностью этого метода является то, что он зарождался не в лабораториях научных институтов, а исключительно сообществом специалистов энтузиастов и добровольцев больных, которые потеряли надежды лечения в официальной медицине и испытали этот метод на себе. Большинство из них добились такого положительного эффекта, которого невозможно достичь всеми современными методами. Особенность метода заключается в том, что он действует не на химическом или физическом уровне клеток, а действует адресно на истинные механизмы перерождения на клеточном уровне – основной причины множества хронических и онкологических заболеваний. Оказывается первичные причины перерождений клеток связаны с нарушениями механизмов регулировки клеток, которые находятся не в ядре клетке, а на многочисленных сенсорных структурах клетки, так называемых интегральных белках ответственных за целостность электрического контура. Глобальная, главная ориентировка регулирования у них происходит не химическим путем, а исключительно на голограммное поле клетки, так называемый электромагнитный каркас клетки. Это так называемое генетическое поле присуще только высокодифференцированным клеткам. У онкологических клеток это поле отсутствует и происходит дезинтеграция всех внутренних структур клетки. Автором книги впервые дана новая концепция электромагнитной саморегулировки, самонастройки клетки как сложной многоэтажной системы. На основе этого построено объяснение действия имплозионного поля прибора на повышение уровня интеграции работы всех структур клетки. Аналогов этому нет. Дано объяснение механизма репарации больных клеток и исчезновения опухолевых. Проанализирован опыт излечения самых различных ранее неизлечимых заболеваний и дана новая комплексная методика их лечения новыми методами, которых нет еще в медицине.
Вы можете обратиться с вопросами или за консультацией к Гарбузову Геннадию Алексеевичу на сайте: garbuzov.org.или по адресу: vitauct@yandex.ru или по телефону 8 928 239 13 64

Похожие статьи
Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне