
Элевационные и инволюционные изменения в гипоталамусе
Элевационные и инволюционные изменения в гипоталамусе как причина геронтоса
Выяснено, что у людей среднего возраста с избыточным весом гипоталамус заметно больше, чем у тех, кто имеет нормальный вес, но того же возраста. Также у людей среднего возраста гипоталамус имеет увеличенные размеры (гипертрофия) по сравнению с нормальными размерами у людей более молодого возраста, а в старости он уменьшается. Очевидно всегда существует связь между гипертрофией гипоталамуса и так называемым воспалением гипоталамуса, а также его гиперфункцией, перезреванием. В всех случаях это аномальное состояние. Организму они не выгодны. Эти процессы находятся за пределами оптимума функционирования системы или органа и при продолжении ведут к его истощению, ослаблению. Подчеркиваю, что речь идёт именно о воспалении мозга, а не клеточном воспалении или на уровне иммунных реакций. Показано, что при хроническом переедании, питании без значительных перерывов в первую очередь страдает гипоталамус. Помочь в этом случае может интервальное питание с ограничением калорий.
«Изнашивание» = перезревание гипоталамуса, в том числе и гипоталамическая возрастная инволюция, онтогенетические трансформации могут происходить от многочисленных причин, в том числе при хронических стрессах, болезнях… Преждевременный климакс и преждевременное старение тоже обеспечиваются через механизмы гипоталамуса.
Продление жизни и неотения (удлинение периода молодости) тоже опосредуются через гипоталамус. Гипоталамус можно расценивать как водитель ритмов = пейсмейкер, где самый большой ритм – длительность жизни. Но изнашивается и стареет гипоталамус и от состояния гормональной перезрелости, избыточности, затем и климакса. Зрелость и перезрелость пролог к старению, а точнее неизбежные последовательные этапы онтогенеза адальтус → матура → сенилит. Одно ведёт к наступлению другого. Климакс при этом следует рассматривать как одно из проявлений резистентности, нечувствительности данного регулирующего органа к различным сигналам нервной системы, к гормонам, нейромедиаторам и др. факторам. Это проявляется как этап истощения, исчерпания и выход за пределы возможности после длительной гиперфункции. После длительной гиперфункции всегда наступает откат в виде гипофункции; после гипертрофии → атрофия. Это приобретение возрастной нечувствительности, толерантности. Регулирующая система при этом выходит за пределы своего оптимума, то есть за пределы коридора гормезиса и работает не на полную мощь, а в итоге вразнос.
Движущие силы онтогенетических изменений в гипоталамусе можно разделить на два уровня:
- Нисходящие процессы с верхних этажей пирамиды систем организма, в частности верховного регулятора, в котором происходит вынужденная элевация (наращивание мощи, гиперфункции, гипертрофии, перезрелости) механизмов верховных гормональных регулировок. Они ведут к гипертрофии и гиперфункции.
- Восходящие с клеточного уровня, которые обусловлены уменьшением количества и ослаблением возможностей стволовых клеток и экзосомальных миРНК с превалированием клеток прогениторного типа, то есть проявления феномена так называемого соматического доминирования, когда состояние ткани и её потребности в репарации определяют не стволовые, а соматические клетки, что, очевидно, связано с лимитом Хейфлика, то есть репликативной ограниченностью клеток. Они ведут к гипофункции и атрофии.
Таким образом, можно утверждать, что гипоталамус стареет в два этапа:
1-й этап гиперфункций – ведёт к адальтусу* и матуре*;
2-й этап гипофункций – ведёт к сенесценту* + сенилиту* = геронтос*.
Первый этап сопряжен с элевационными процессами по Дильману. Второй этап сопряжен с угасанием, инволюцией органа по Гращенкову. Таким образом, по Гарбузову геронтос – это двухэтапный процесс старения от элевации до инволюции. Кроме того, геронтос – двусторонний процесс: от сенилита до сенесцента. Особенностью данной гипотезы является то, что в процесс общего старения включен еще и сенесцент. Таким образом, геронтос - это единство сенесцента и сенилита, то есть взаимоувязанных процессов.
При этом Гарбузов утверждает, что сенесцент это проявление, следствие онтофизиса*, а не формального проявления лимита Хейфлика, феномен который связывали с укорочением теломер. Причем сенесцент и онтофизис являются следствием эпигенетических перестроек, в отличие от всех остальных этапов онтогенеза, которые являются программированным морфогенезом.
Такое наращивание происходит из-за постепенного приобретения резистентности и толерантности в данной системе с обратными связями: верховный регулятор – железы и органы мишени. С возрастом большое количество функций ослабевает из-за появления в системе звеньев с резистентностью, они попросту недорабатывают, ослабевают. Резко возрастают на стадии зрелости = матуры количество половых стероидов как тестостерон, эстрогены, а затем их количество идёт на убыль. С другой стороны уменьшается количество выработки мелатонина, гормона роста - соматропина и др. Меняется баланс нейромедиаторов, в частности увеличивается, а затем уменьшается кортизол, снижается ацетилхолин и т.д. Вся система регулировок сдвигается, причем не каждый отдел сдвигается сам по себе по своим правилам, а в соподчинении с другими параметрами гормонов. Такие вторичные программы и механизмы соподчинения предусмотрены генетически в филогенезе. Весь этот комплекс разноплановых систем регулировок, которые движутся в разных направлениях, есть проявление общего сдвига регулировок.
Клеточное старение (сенесцент) как проявление клеточной нечувствительности, неуправляемости и перехода на эпигенетическое регулирование, когда оно становится доминантным началом в жизненных циклах клетки
Напомню, что сутью сенесцентных клеток является как репликативное торможение, так и усиление в них нечувствительности, толерантности во многих сигналинговых путях и рецепторных структурах, что ведёт клетки к рецессии, то есть снижению их как функциональной работоспособности, так и активность стволовых клеток. Такие клеточные системы начинают вырождаться. Подробно об этом в моей книге: «ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ». Для того чтобы объяснить природу сенесцента клеток, следует предварительно рассмотреть как клетки входят в состояние стресса, а затем в состояние сверхадаптации. Речь пойдет именно о клеточном стрессе, а не общеорганизменном, известного под названием Общий Адаптационный Синдром (ОАС), который описал Г. Селье. Факторов клеточного стресса бывает множество, но механизмы их стресса и итоговые последствия тоже универсальны, сходны. Автором предложено их обозначить как Универсальный Адаптационный Клеточный Ответ (УАКО). Он может быть условным, то есть вызванным внешними факторами, так и безусловным, то есть врожденным, связанным с внутренними факторами онтогенеза и онтофизиса. В связи с этим сенесцент следует рассматривать тоже как следствие внутренних факторов условий развития. В разделе: Гормезиорез* и гомеорез* – причина сенесцента, сенилита через эпигенез и деградацию белков показано, что причиной клеточного хронического стресса = сенесцента может быть как выход организма в целом за пределы гормезиса, связанный с возрастом, так и вычленение и содержание клеток вне организма в условиях культуры ткани, когда всё это выводит на длительное функционирование клеток за пределами гомеостаза. В итоге жизнедеятельность таких клеток начинает выходить за пределы оптимума. Как результат снижается ВИТАУКТ* организма и Жизненная Сила клеток. Механизмы клеточного стресса реализуются через образование белков теплового шока (БТШ), которые запускают белки-сиртуины, в частности SIRT1, вслед за которыми начинаются эпигенетические перестройки со многими последствиями. Любой стресс активирует клеточные функции, делает их более чувствительными за счёт механизмов УАКО*. АОС начинается с активации надпочечников и усиливает выработку ими гормонов стресса адреналина, кортизола, глюкокортикоидов, обладающих способностью стабилизировать мембраны, другие гормоны работают как экспрессоры для генов. Клетка приобретает повышенную сопротивляемость к негативным факторам, функциональность, но при этом может выйти за пределы гомеостазов, а значит работает на износ. К внутренним негативным факторам можно относить и онтогенетический ответ клеток на возрастные изменения в организме. Причем в отличие от более ранних этапов онтогенеза, вплоть до юности, они сопровождаются генетическими регулировками, включая механизмы морфогенеза, например пубертатное оволоснение. Но в последующем динамика управления онтогенезом с переходами на этапы адальтуса, матуры, сенилита переходит на превалирование регулирования на уровне эпигенеза. Эпигенез становится доминантным фактором, «жокеем» дальнейшего развития с целым шлейфом различных анизомалий. Он же эпигенез становится и ограничительным фактором, обуславливающим мортус, то есть феноптоз*. Этим тип феноптоза человека отличается от рыбы горбуши, причиной смерти которой является выраженное гипергипоталамическое доминирование, ведущего к избытку репродуктивных гормонов с многочисленными последствиями не совместимыми с жизнью.
В любом случае это режим в одних условиях для дегенеративных перерождений, утрате тканью дифференциальных функций, а в других – путь к сенесценту или онкогенезу. Особенность клеток животных в том, что к сенесценту клетки изначально предрасположены, настроены на это. Но это не программативная настроенность, а коммитативная – то есть на уровне эпигенеза – многогранный комплекс изменений происходящий, как в состоянии стресса, так и онтофизиса.
Как гипоталамус будучи гетерогенной, многофункциональной структурой определяет сенилит
За что отвечают ядра гипоталамуса?
В ядрах гипоталамуса происходит тончайшая координация деятельности вегетативной нервной системы, которая управляет всеми внутренними органами, регулирует процессы обмена веществ в организме.
Благодаря четкой и слаженной работе различных отделов гипоталамуса сохраняется относительная устойчивость различных функций организма, что совершенно необходимо для его нормального существования.
За что отвечает гипофиз и гипоталамус?
Гипоталамо-гипофизарная система регулирует такие важные функции организма, как температура тела, сон, аппетит, обмен веществ, половое развитие, репродуктивную функцию и многое другое. Она также контролирует выработку таких гормонов, как гормон роста, пролактин, АКТГ, тиреотропин, лейтенизирующий гормон и фолликулостимулирующий гормон.
Гипоталамус. Характеристика основных ядерных групп. Участие гипоталамуса в регуляции вегетативных функций и в формировании эмоций и мотиваций
Группа передних ядер. Это супраоптическое, супрахиазматическое, паравентрикулярное и др.
Средняя группа ядер. Это вентромедиальное, дорсомедиальное и др.
Наружная группа ядер. Образована латеральным гипоталамическим ядром, ядром серого бугра и т.д.
Задняя группа ядер. Состоит из заднего гипоталамического, перифорникального, премамиллярного, медиального мамиллярного и латерального мамиллярного ядер и др.
Особенностью гипоталамуса является то, что его ядра могут возбуждаться двумя способами. Первый способ – это нервный путь. Второй способ – это возбуждение ядер гипоталамуса гуморальными влияниями. Данная особенность гипоталамуса связана с наличием в гипоталамусе 3 типов нейронов. Первый – нейросекреторный. Второй тип нейронов – ретикулярный. Третий тип нейронов – это слабоветвящиеся нейроны с короткими дендритами.
Связи гипоталамуса. Он имеет хорошо развитые афферентные и эфферентные пути. Он получает афферентные пути из коры больших полушарий мозга, из таламических ядер, ядер базальных ганглиев, ретикулярной формации среднего мозга.
Ядра переднего гипоталамуса – супраоптическое и паравентрикулярное – связаны системой волокон с гипофизом. Эти волокна служат не только для проведения импульсов, но и для транспорта продуктов нейросекреции, которые вырабатываются нейронами этих ядер (вазопрессин, окситоцин, статины, либерины).
В составе гипоталамуса находятся нейроны, вырабатывающие гормоны или специальные вещества, которые в дальнейшем, действуя на клетки соответствующих эндокринных желез, приводят к выделению или прекращению выделения гормонов (так называемые рилизинг-факторы). Все эти вещества вырабатываются в нейронах гипоталамуса, затем транспортируются по их аксонам в гипофиз. Ядра гипоталамуса связаны с гипофизом гипоталамо-гипофизарным трактом, который состоит примерно из 200 000 волокон. Свойство нейронов вырабатывать специальные белковые секреты и затем их транспортировать для выброса в кровяное русло называется нейрокринией.
Гипоталамус является частью промежуточного мозга и одновременно эндокринным органом. В определенных его участках осуществляется трансформация нервных импульсов в эндокринный процесс. Крупные нейроны переднего гипоталамуса образуют вазопрессин (супраоптическое ядро) и окситоцин (паравентрикулярное ядро). В других областях гипоталамуса образуются рилизинг-факторы. Одни из этих факторов играют роль гипофизарных стимуляторов (либерины), другие — ингибиторов (статины). В дополнение к тем нейронам, аксоны которых проецируются в гипофиз или в портальную систему гипофиза, другие нейроны этого же ядра отдают аксоны в многие участки головного мозга. Таким образом, один и тот же гипоталамический нейропептид может выполнять роль нейрогормона и медиатора или модулятора синаптической передачи.
Велика роль в регуляции и координации функций вегетативной нервной системы. В регуляции функции её симпатической части участвуют ядра задней области гипоталамуса, а функции парасимпатической части вегетативной нервной системы регулируют ядра его передней и средней областей. Стимуляция передней и средней областей вызывает реакции, характерные для парасимпатической нервной системы — урежение сердцебиений, усиление перистальтики кишечника, повышение тонуса мочевого пузыря и др., а раздражение задней области проявляется усилением симпатических реакций — учащением сердцебиений и т.д.
С состоянием вегетативной нервной системы тесно связаны вазомоторные реакции гипоталамического происхождения. Различные виды артериальной гипертензии, развивающиеся после стимуляции Г., обусловлены комбинированным влиянием симпатической части вегетативной нервной системы и выделением адреналина надпочечниками, хотя в данном случае нельзя исключить влияние нейрогипофиза, особенно в генезе устойчивой артериальной гипертензии. Таким образом, возрастную гипертонию, хроническую усталость, бессонницу… можно рассматривать не только как примеры разболтанности, изношенности, разрегулировки неврологической стороны деятельности гипталамуса, но и как возрастных неизбежных его перестроек, связанных с онтогенезом. Тогда как поседение волос, ухудшение зрения, ухудшение качества кожи, морщины на ней, мышечная саркопения, атеросклероз, диабет… – проявления трофической функции гипоталамуса в целом и в частности его ядер определяющих это. Гипоталамус стареет гетерохронно и гетерогенно. Все многочисленные ядра являются представительствами многочисленных функций регулировок, за которые они отвечают. У каждой функции, системы имеется своё ядро-представительство в Генштабе. Деятельность всех этих представительств согласована. С перестройками в одних системах могут происходить подстройки в других. Обобщенно - весь этот спектр возрастных изменений деятельности гипоталамуса – это многочисленные проявления выхода за границы коридора Гормезиса, когда происходит потеря многочисленных сил Витаукта – интегральной Жизненной Силы. Обобщенно - Витаукт это сила определяющая слаженный интегрированный оптимум работы всех систем. Это сила, а точнее программа гармонизирующая работу всего Генштаба. Витаукт состоит из многочисленных ручейков автоматической саморегулировки, когда их оркестр играет одну симфонию под названием Гармония.
Итак, гипоталамус гетерогенная структура, развивающаяся гетерохронно и, тем не менее, определяющая специфику сенилита, стадий онтогенеза. Скорость онтогенеза определяют не все ядра гипоталамуса, а только те, которые определяют репродуктивную зрелость и трофику тканей.
К числу гипоталамических ядер, ответственных за реализацию программы старения, предположительно относят супрахиазмальное ядро и срединную группу ядер, включая аркуатное, вентромедиальное и дорсомедиальное ядра. Процесс старения в гипоталамусе сопровождается:
- нарушением баланса возбуждение/торможение за счет дисрегуляции ГАМК-ергической системы,
- уменьшением частоты импульсации нейронов,
- нарушением обмена кальция, магния,
- увеличением продукции оксида азота,
- снижением аутофагии,
- нарастанием нечувствительности, рефрактерности к сигналам (невосприимчивости), толерантности,
- усилением првоспалительных процессов,
- оксидативный и нитрозативный стресс,
- снижением уровня эргичности клеток, в т.ч. выработки АТФ,
- нарушением процессов внутриклеточного сигналинга, в частности Sirt1, mTOR, NF-κB, AMPK, P53, FoxO…
- …
Нейроны ДЯ и ДМЯ содержат медиаторы глутамат и ГАМК, при этом доля последнего преобладает. В ВМЯ преобладают глутаматергические нейроны, а доля ГАМК-ергических невелика. При старении у крыс усиливаются взаимно противоположные процессы возбуждения и торможения, по-видимому, нарушая баланс ГАМКергической и глутаматергической систем.
Также важная роль в гипоталамическом старении принадлежит таницитам, что выражается в нарушении их связи с нейронами и барьерной функции. Стволовые клетки гипоталамуса, расположенные преимущественно в области срединной группы ядер, выделяют экзосомальные миРНК, которые также ответственны за антивозрастные эффекты этих клеток. При этом количество самих стволовых гипоталамических клеток и экзосомальных миРНК уменьшается по мере старения.
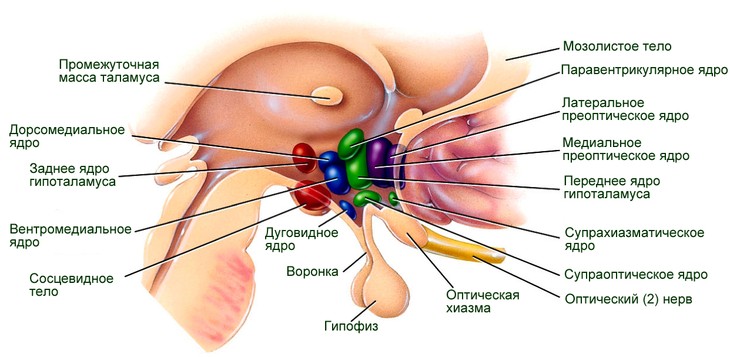
Рис. 1. Многоядерное строение гипоталамуса. Гипоталамус как орган отвечает за многочисленные нейрогормональные функции, в том числе и за созревание организма и его старение.
Важным отличием и особенностью гипоталамуса животных относящихся к долгожителям является наличие в гипоталамусе демфинговых структур (амортизаторных), механизмов, которые сглаживают провоспалительные процессы на уровне клеток, тем самым пролонгируя функционал этих клеток. Более мощные и самоомолаживающиеся гипоталамусы обеспечивают более продвинутые сроки продолжительности жизни.
Способность к самоомолаживанию гипоталамуса обеспечена на коммитационном уровне. Животные долгожители коммитированы на мощное самоомоложение, реювенилизацию клеток гипоталамуса, преодоление в них сенесцента. Это аналогично тому как жестко регулируется уровнем гормонов и их соотношением продолжительность жизни 1- или 2-х летних растений, в отличие от их антиподов многолетников. Уровень гормонов реювенилизаторов – цитокининов у короткоживущих растений практически отсутствует, не предусмотрен, тогда как у долгоживущих такая возможность выработки реювенилизаторов постоянно открыта, что обеспечивает закладку новых почек для роста. В свою очередь у короткоживущих животных не предусмотрены демферные механизмы для сдерживания и противодействия провоспалению. Регулировка происходит через посредство главного апекса – аналога гипофиза. Продолжительность жизни короткоживущих растений коммитирована специальными механизмами. К такому же типу регулировок продолжительности жизни относится и рыба горбуша, которой строго предписана монокарпическая жизнь, то есть отмирание после первого нереста в 8 лет. Этот тип регулировки онтогенеза животных определяется как гипергипоталамическое доминирование, а у растений 1- и 2-х леток как гиперапикальное доминирование.
Термины
- Адальтус – этап онтогенеза после юности, то есть генеративной зрелости.
- Геронтос – это единство сенесцент ![]() сенилит. Общее старение которое исходит как со стороны сенилита, так и со стороны сенесцента.
сенилит. Общее старение которое исходит как со стороны сенилита, так и со стороны сенесцента.
- Гормезиорез - состояние, когда параметры устойчивого гормезиса претерпевают возрастные изменения. Термин предложен Г. Гарбузовым.
- Гомеорез - состоит из слов «гомео» — постоянство и «рез» — смещение, сдвиг. Состояние, когда устойчивый гомеостаз может перейти в системе на новый уровень стабильности, констант.
- Матура – этап онтогенеза, перезрелости, с выходом за пределы, границы оптимума коридора гормезиса, предшествующий сенилиту.
- Онтофизис - термин предложен Гарбузовым Г. А. – подразумевает у растений не простое закрепление у клонов неких особенностей роста и плодоношения, а проявление более общего процесса, то есть сохранения степени онтостадийной зрелости, включая высшую степень онтостадийности, когда полностью заблокирован рост или плодоношение на нижних побегах. В отличие от высшей степени онтофизиса у побегов растений в животном мире аналогом этому является сенесцент у клеток. Онтофизис = сенесценту. Разблокировать такое состояние у растений возможно только при особых условиях реювенилизации, то есть снятии апикального доминирования и обязательном превалировании цитокининового фона. При этом расширено понимание этого явления не только для клонов, но и в пределах данной особи растения, когда новое поколение побегов в пределах одного и того же дерева все проявляют продвинутую физиологическую или морфологическую стадийную зрелость. Программы на ювенильность у них заблокированы. Если расширять понимание этого термина на животных, то подразумевается закрепление неких возрастных и онтогенетических перестроек, которые сохраняются и у линий клеток в культуре ткани, аналогично тому, как это сохранение происходит и у клоновых потомств в виде топофизиса (особенностей роста: ортотропного или плагиотропного) или циклофизиса (способность к закладке или не закладке тех или иных генеративных органов), или онтофизиса (с сохранением темпов клеточного роста или самообновления или их полной блокировки). Особенностью клеток животного типа является то, что до сих пор не найден гормон реювенилизации сенесцентных или онтофизисных клеток, что и является причиной «непреодолимости» лимита Хейфлика. При этом особенностью выращивания клеток в культуре ткани является быстрый «перескок» через все онтостадии и остановке на крайней онтофизисной стадии.
- Сенилит – старение, которое зарождается на верхнем общеорганизменном уровне.
- Сенесцент - репликативное старение, которое зарождается на уровне клеток.
- Феноптоз - тоже что и клеточный апоптоз, то есть запрограммированная смерть клеток, только на уровне всего организма.

Похожие статьи
Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне