
Эпигенетические механизмы молодости и старения клеток
Статья - глава из книги Гарбузова Г А: ЭПИГЕНЕТИКА МОЛОДОСТИ
До сих пор нет научного ответа откуда берёт начала сенесцент* (клеточное старение) и что является его драйвером? Для чего он нужен в мире животных и почему мир растений обходится без него? Существует множество разрозненных концепций, пытающихся дать ответ на разных уровнях, но нет единой всеобъемлющей теории, которая могла бы дать такой ответ, который бы позволял осознать необходимые приёмы для преодоления сенесцента. Сплошные концепции, разрозненные пазлы…, не дающие воссоздать всю многогранную и многоэтажную картину проблемы. Не хватает множество звеньев, одним из которых является нерешенная проблема сенесцента на уровне взаимодействия хроматина и его эпигенома…
Хроматин с позиций БИОКРИСТАЛЛа и что общего с «Магическим» кристаллом?
Автор считает, что некоторые стороны активности хроматина объяснить односторонне с позиций или биологии, химии или физики невозможно, поэтому предлагает рассмотреть свою концепцию функций хроматина не просто с позиций единства этих свойств (супербиокристалл), а то что определяет его квази, то есть за пределами этих возможностей (феноменами), но кроме того активно реагирующего на все сигналы, причём способного к самовосстановлению после прекращения воздействия внешних факторов.
Что такое эпигенетическая мантия? Хроматин создаёт вокруг себя эпигенетическую мантию (вместе они - единство противоположностей) и при этом каждый из них проявляет сверхвысокую чувствительность ко всем изменениям с противоположной стороны, а также каждый из них определяет статус противоположной стороны.
При этом мантия (= эпигеном) состоит не просто из гистоновой рубашки, но и других «слоёв», которые образованы из метильных (метилом), ацетильных и других групп (которые, очевидно, определяют скопление тех или иных электростатических зарядов на стенке ядра, меняют степень и состав ионизации), pH-среды, а также окружающей среды, кариоплазмы (с её переменным спектром компонентов), которые в совокупности представляют собой те или иные эдафотопы*. Напомню, что в экологии эдафотопы означают факторы среды, которые определяют состав сукцессий (живых сообществ). В нашем понимании эдафотопы определяют свойства мантии и эпигенома. Получается, что хроматин и его мантия это многосферная структура. Они работают как одно единое целое. Поэтому, часть поступающих внешних сигналов к хроматину трансформируется через мантию. Все онтогенетические сдвиги фиксируются первично на мантии, приводя в ней к ряду изменений. В первую очередь это происходит в областях генома над программами ответственными за проявление той или иной степени (уровня) потентности стволовых клеток. Это приводит к глушению или активации этих зон (их «видимость» или «невидимость»). Можно говорить о мантии ювентального типа и мантии сенесцентного типа.
Возможно, на степень прозрачности эпигенома влияет pH среды клетки, которая у сенесцентных и онкологических клеток всегда закислена, а энергетика их всегда ближе к гликолизной; у которых наблюдается изменение активности фермента β-галактозидазы и обусловлена увеличением количества этого фермента в лизосомах и цитоплазме, а также увеличением числа самих лизосом в клетке.
Очевидно, гены, определяющие степень стволовости выделены в отдельный блок на отдельной хромосоме, и являются центральным блоком управления. Гены стволовости размещены последовательно: пока не отработают-запустятся гены первого порядка (тотипотентные) не запускаются гены второго порядка и так по цепочке. Это принцип первичного последовательного разворачивания, трансформинга. Вместе с разворачиванием происходит и последовательное «глушение» генов высших порядков потентности. Отработавшие гены 1-го порядка запускают работу генов 2-го порядка (плюрипотентных) и т.д. В организме имеются ткани и 16 систем. В этом случае в каждой из них предшествующий уровень потентности отключен, но программа морфогенеза в каждом типе данной ткани запускает свой базовый уровень потентности и последующих нужных уровней потентности для своего набора типов стволовых клеток.
Итак, если порядок стволовости клеток передаётся в большей степени последовательно сквозь любые потомства, то морфогенетические признаки распределяются мозаично, только в пределах данной ткани. Стволовость накладывается на морфогенез…
Такой блок регулировок переключения потентностей должен работать соподчинённо и в унисон-синхронно, для чего должен быть выделен в отдельную группу, так как работает по другим правилам и задаётся статусами мантии над этими блоками генов. Механизм регулировок здесь начинается не на уровне генов, а на уровне эпигенома, а точнее в целом всей мантии, и осуществляется на других этажах хроматиново-эпигеномных взаимоотношений, то есть на других принципах.
Такая высочайшая сложность иерархий систем определяет необходимость создания двухэтажных систем регулировок. В таких гиперсложных иерархических конструкциях организм, по сути, приобретает свойства «квазигосударства», («голого», без системы надлежащего управления), то есть фактически не управляемого только через верхние общесистемные уровни. В таких условиях возникает необходимость более точных регулировок на местах. Этим можно объяснить необходимость возникновения в животном мире сенесцентного и иммунного типа клеток. Так возникают дополнительные системные автономии управления на уровне клеток. Это позволяет всю эту 3D конструкцию рендерировать* (перевести, упростить) в 2D плоскость с помощью биокристалла. Сигналы преобразуются из специфических в неспецифические, универсальные. Из множества разрозненных сигналов остаётся всего лишь несколько, на которые геном отвечает стандартно.
Суперпозиционная регулировка мантией генома. Ювентальные или сенесцентные типы мантии – это уже различные её состояния, суперпозиции. Суперпозиция – это когда один и тот же объект может находиться в различных состояниях. Именно через эти локальные суперпозиции в мантии происходит «блокировка» или «активация» частей генома, находящегося под ними (прозрачность – непрозрачность). Это аналогично тому как если направить поляризованный свет через горизонтальный фильтр. Если на него направить свет, уже поляризованный горизонтально, он пройдёт без проблем, но в случае если направить свет, поляризованный вертикально, он будет полностью заблокирован. А вот если направить неполяризованный свет, он пройдёт лишь частично, поскольку в нём присутствуют колебания во всех плоскостях, включая горизонтальную. Это означает, что мантия – это своеобразный фильтр для сигналов. Главный параметр (поляризатор) этого фильтра – это степень метилирования, которое в первую очередь отражается над областью стволовых клеток.
Мантия может быть поляризована на ювентальность и сенесцентность (степень метилирования общая и локальная…). От статуса состояния этого фильтра зависит пройдут ли те или иные сигналы к геному или нет. Причём состояние этого фильтра определяют эдафотопы. При этом фильтр-мантия сам себя адаптирует, преобразует, поляризует к изменяющейся среде. Это и является основой для устойчивых преобразований в мантии и смене её статуса. Мантия у сенесцентных клеток иная чем у ювентальных. Очевидно она закислена, протонирована, так как клетки ближе к гликолизным, а значит менее поляризована минусовыми электрозарядами и сильнее обеспечивает стягивание (натяжение) хроматина, что делает его невидимым. Это уже иной суперпозиционный статус фильтра, то есть с закреплёнными свойствами, а не переменный, ситуационный. Как следствие меняются при этом и эдафотопы (среда), которые определяют статус мантии. Закреплённые свойства означает стабильную смену активности ряда стволовых клеток. Это в свою очередь закрепляется в новых фенотипах и модификатах клеток… Их нельзя путать с морфотипами! Происхождение их иное. Со сменой активности ряда генов (стволовых прогениторов) происходит смена эдафотопов, которые сами становятся обусловленными структурными изменениями на уровне систем.
Взаимодействие между эдафотопами, мантией, эпигеномом, хроматином. Доказательством того, что эпигеном, а точнее мантия (в которую он входит) не просто некий буфер для генома, а активный участник регулировок, и который сам подвергается различным влияниям и регулировкам могут быть многочисленные факты. Таковыми могут быть антиоксиданты, которые своё действие оказывают часто не напрямую на геном, а через эпигеномную мантию. Так примерами могут быть флавонолы какао, куркумин, которые действуют как регуляторы и противодействуют провоспалению, одного из маркеров сенесцента. Гася провоспаление, флавонолы могут сдерживать сенесцент. Основная гипотеза, подтверждённая рядом исследований, заключается в их способности ингибировать ядерный фактор «каппа-Б» (NF-kB). Это главный молекулярный «переключатель», активирующий гены воспаления в ответ на стресс и повреждения. Причём, доказано, что факторы стресса и антистресса могут действовать сначала на уровне цитоплазмы, а она (является частью эпигеномно-мантийно-хроматиновой частью биокристалла) опосредует на уровне эпигенетических механизмов, в частности через совокупность специфических меток, определяющих активность генов. Это означает, что через мантийно-эпигеномно-хроматиновый комплекс (биокристалл) могут действовать факторы генеративного созревания организма, смены в нём различных профилей, в том числе и гормональный, иммунный и другие… С позиций действия их на каждую сторону биокристалла по отдельности они могут вызывать специфические ответы, но если рассматривать этот комплекс как единый-неделимый, то проявится неспецифический ответ. Благодаря универсальности ответов этого комплекса, работающего на принципах «биокристалла» (напомню, что суперкристаллы отвечают на разные энтропийные факторы единым ответом), всё их разнообразие вызывает общий ответ, приводит к общему знаменателю, действует по одному сценарию, неспецифически, то есть усиливая сенесцент или ему противодействуя. Итак, можно утверждать, что специфический ответ всегда проявляется через геном, тогда как неспецифический всегда проявляется через статус хроматина.
Внешняя среда клеток (интерстиций) меняет и их внутреннюю среду, то есть внутриклеточные эдафотопы как в цитоплазме, так и в кариоплазме. Как результат идёт подстройка состояния хроматина (прозрачность – непрозрачность) и активности эпигенома и отчасти генома. Это подтверждает идею, что сенилит* тоже соучастник сенесцента, ускоряет его развитие, меняет качество хроматина сверху на системном уровне.
В таком случае важен ответ: когда происходит ответ этого комплекса специфический, а когда неспецифический? Для этого есть система распознавания. Ключами для распознавания являются провоспалительные сигналы и генные провоспалительные ответы. Сенесцент держится на провоспалительных механизмах, которые состоят из ключа (фактора) и генного ответа (замка). Только в обсуждаемой нами системе ключей для вскрытия провоспаления может быть множество, а замок реагирующий на них общий – биокристалл, то есть вся его многокомпонентная конструкция. Определённое сочетание его компонентов (сложный код замка) позволяет ему реагировать на ключи, точнее на спектр ключей. Замок реагирует по принципу: прозрачный – непрозрачный участок хроматина. Как сукцессии зависят от эдафотопа, так и мантия зависит от среды. По отдельности эти компоненты не работают, а только в составе биокристалла. Замок – это интегрированная конструкция (очевидно с помощью конформационных белков - КБ).
Конформационные белки* зависят от ряда факторов и могут участвовать в разрыхлении хроматина и увеличении его прозрачности? Все вместе они представляют собой сенсорный дисплей, который срабатывает только при сочетании синхронности всех его компонентов. Таким образом, для запуска сенесцента нужны многофакторные условия, создающие готовность замка к ответу.
Белки теплового шока* (HSP = БТШ) и сенесцентные клетки связаны через процесс старения организма (сенилит), который регулируется на верхнем системном уровне. Так происходит взаимопроникновение, взаимообусловливание сенесцента и сенилита. БТШ тоже действуют через систему провоспалений, они тоже могут усиливать воспаление. Действие БТШ может быть двойственное: как анти-, так и провоспалительное. Всё зависит от их уровня так и чувствительности к ним. Очевидно система конформационных белков сенесцента и БТШ сенилита – это единая система самонастраивания организма как единого целого (интегративная функция). Сенилитное провоспалительное действие организуется через БТШ посредством иммунной системы, провоспалительная реализация сенесцента - через Биокристалл клеток. Любой этаж старения реализуется через провоспаления. Единый уровень провоспалений геронтоса* регулируется через провоспаления сенесцента и сенилита, которые в свою очередь регулируются на уровне биокристалла и иммунной системы. Тогда по аналогии ювентос* = ювентальность + ювенил, где в любом из них отсутствует провоспаление. Через единство (доминирование) геронтоса (сенесцента и сенилита) и ювентоса (ювентальность + ювенил), очевидно, регулируется длина Онтодианной Линии Жизни.
Итак, если есть замок (биокристалл), то к нему всегда есть и отмычка.
Авторский концепт о природе онкологии. Автор считает, что онкоклетки – это всего лишь противоположная крайность процесса сенесцента, так как они схожи по гликолизности, провоспалениям, митохондриальной недостаточности и по эпигенетической природе происхождения. На начальных этапах появления онкологических клеток нет мутаций как таковых, и только на продвинутых этапах их развития эпигенетическая их природа доходит до крайности, когда они не могут поддерживать свои прежние гомеостазы и начинаются вторично транспозонные сдвиги на геноме. То есть онкопроцесс начинается как модификат сенесцента на одних и тех же «рельсах» в эпигенезе, различия лишь в степени зашкаливания выхода за пределы гомеостазов и уровня провоспаления: в одних случаях клетка уходит в сенесцент, в других - в онкологический модификат. Сенесцент и малигнизация – это различные модификаты одного и того же процесса. Напомню, что согласно концепции Биокристалла – эпигеном и хроматин – это взаимовлияющие друг на друга стороны одной системы и при стрессах этот кристалл реагирует изменениями, но в определённых ситуациях процесс напряженности взаимоотношений их может стать критическим, за пределами саморегулировки гомеостазов и обратного исходного возращения, когда в свои суперпозиции кристалл не возвращается. Подобие чрезмерного растяжения резины, которая имеет предел натяжимости, а далее идёт деформация… При онкологии локальная запредельная деформация хроматинового биокристалла глубже, что делает невидимыми гены регулировок репликаций, как результат репликативная резистенция, неуправляемость, нерегулируемость… Если рак осуществляется полностью на принципах энтропогенеза (энтропия = саморазрушение), то сенесцент – это организуемый организмом процесс. Различия между сенесцентом и раком в степени проявления провоспаления и глубине гликолиза, а также тем, что рак развивается локально, причём это возможно даже в условиях тотального сенесцента организма, где сенесцент разносится с помощью иммунитета, тогда как раковые клетки не поддерживаются иммунной системой. Таким образом, нельзя утверждать, что сенесцент является защитой, преградой для рака. Это приводит к геномной нестабильности. Затем инсерции транспозонов могут повреждать гены, вызывать хромосомные перестройки, нечто подобное мутациям, но это не мутационная природа опухолей. Напомню, что у растений нет настоящих раковых клеток и сенесцентных. Есть только ложное подобие сенесцента (ложноподобное старение клеток, так как сами они не стареют), которое реализуется через онтофизис*. Это тоже указывает на то, что у сенесцента и онкологии общие начала.
Принципы и механизмы смены возрастного статуса клеток. Такие всеобщие изменения (турбулентность) приводят к перенастройкам разных уровней клетки друг к другу, самоуравновешиванию, но уже на новом уровне, смене балансов устойчивости, смене гомеостазов. Эти новые гомеостазы стабилизируются уже не только функционально, но и структурно.
Через новые гомеостазы меняются эдафотопы и система переходит из турбулентного зыбкого состояния в ламинарное, то есть в устоявшуюся позицию (но новую): одно обуславливает другое.
Универсальность механизмов сенесцентирования во всех морфотипах тканей. Такая турбулентность и переход на новую суперпозицию и новый гомеостаз может происходить над каждым паттерном, представляющим собой тот иной тип стволовых клеток в разных типах тканей. Ткани разные, а степень потентности стволовых различных клеток может быть одинаковой. То есть на морфогенетические процессы накладываются онтогенетические (эпигенез). Таким образом, морфотипы могут быть разные, а возрастные эпигенотипы одинаковые. Но возможно в разных случаях и наоборот: эпигены разные, а морфотипы одинаковые, что может привести к дополнительным проявлениям «надморфотипов». Это один из механизмов внешнего проявления сенилита: старческая кожа… Смена эдафотопов является условием для «закрепления» свойств = статуса мантии, а мантия в свою очередь меняет статус генома. Таким образом, становятся возможными проходы перестроек через все уровни систем организма и на всех этапах Онтодианной Линии Жизни.
Статусы или типы-суперпозиций мантии и определяют особенности транскрипции генома.
Почему клетки растений не стареют, у них нет сенесцента? Важно понять почему тогда у растений, у которых тоже есть тоже эпигеном и его метилирование, но нет сенесцента? Статус тканей здесь определяется только через онтофизис*. Все настройки идут только сверху, на уровне систем, но нет коммитирования с клеточного уровня! Коммитация начинается на уровне мантии. Значит это не генетическая программа. Организация регулировок у растений проще – одноэтажная, тогда как в животном мире – она двухэтажная. Организация строения организма животного на много сложнее (уровень квазисистем), чем у растений, требуется соблюдать множество условий-ограничений, а значит управлять всей этой архитектурой требуется более точно, для этого и задействован механизм сенесцента в качестве важнейшего ограничителя.
Авторская концепция механизмов регуляции становления сенесцента в клетке. Геном это всего лишь «клавишная партитура», исполнитель, где дирижером является мантия. По сути, мантия – это сенсорное поле, определяющее правила игры. Это не только «химия», но и магнитоэлектростатический регулятор-буфер. Поэтому мантия может активно влиять на хроматин, открывая или закрывая его генные локусы. Похоже благодаря некоторым свойствам мантия в ряде мест, то есть локально, может быть «прозрачной» или «непрозрачной» для внешних сигналов. Степень метилирования или ацетилирования определяют и степень насыщения тех или иных зарядов, или ионов, и активности конформационных белков, что способствует или не способствует транскрипции. Эту «прозрачность» определяют скопления электростатических зарядов в разных её точках (ячейках или витках, или над активными генами…), а это в свою очередь определяет степень метилирования ряда гистоновых групп, которые входят в состав мантии. Определённые «рисунки» метилирования хроматина в разных его локусах, как чертежи обуславливают статус ювентальности или сенесцентности клетки. Таким образом, сенесцентность определяется не только теломерами (как это позиционировалось ранее), а в первую очередь всем комплексом эпипаттернов над всем хроматином. Такой хроматин меняет свои химические и электрофизические свойства, становится более компактизированным, свёрнутым, упакованным и перекрывает гены регуляции репликаций, в частности блок G1/S. Такая клетка становится не только не способной к репликациям, репарациям, но и резистентной, нечувствительной к сигналингу, а значит слабо функциональной. Так появляется механизм реализация и управления сенесцентом.
Кстати, одной из причин сенесцентного блокирования клеток является возможность того что механизмы регенерации имеют общие гены с онкологическими процессами и стимулирование регенерации на фоне провоспалительного статуса легко может запустить онкологию. Таким образом, автор пересматривает ранее устоявшееся мнение, что сенесцент это превентивная защита от онкологии, нет: сенесцент и блокирование высокопотентных стволовых клеток на ранних этапах становления ткани – это защита от регенеративных процессов, которые в условиях организма животного легко бы могли перейти в онкологию. Возможно, доброкачественные опухоли связаны с прогенетирными стволовыми клетками (регулируются преимущественно на уровне эпигенеза), тогда как злокачественные опухоли затрагивают гены более глубоких уровней потентностей. Как вывод, можно утверждать, что мантия и биокристалл связаны не только с эпигеномом, но и геномом. Онкология – это тоже опосредованное эпигенетическое заболевание, но реализуемое на геномном уровне.
Миссия энхансеров. Поддержку («подсвечивают» электрозарядами) этой прозрачности или непрозрачности могут обеспечивать специальные окологенные «вставки» (энхансеры*?) вокруг особо важных функциональных генов, отвечающих за репарацию, степень стволовости клеток… Очевидно, энхансерам помогают конформационные белки: их активность зависит от статуса эдафотопов и имеют две позиции «да» или «нет», включено или выключено. На определённые сигналы эти вставки усиливают внешний заряд и тем самым «открывают прозрачность» или её «закрывают», а точнее разрыхляют или уплотняют тело хроматина. Другими словами, это процесс компактизации (конденсации) хроматина. Степень конденсации хроматина связана с его транскрипционной активностью. Неконденсированный хроматин более транскрипционно-активен, чем... Хроматин свёрнут в виде петель, причём эти петли накручиваются и разворачиваются вокруг оси, создавая объёмное тело из этих петель, где каждая петля состоит из спиралей-витков, а в итоге всё это представляет собой многократное сложение жгута (см. рис. 1).
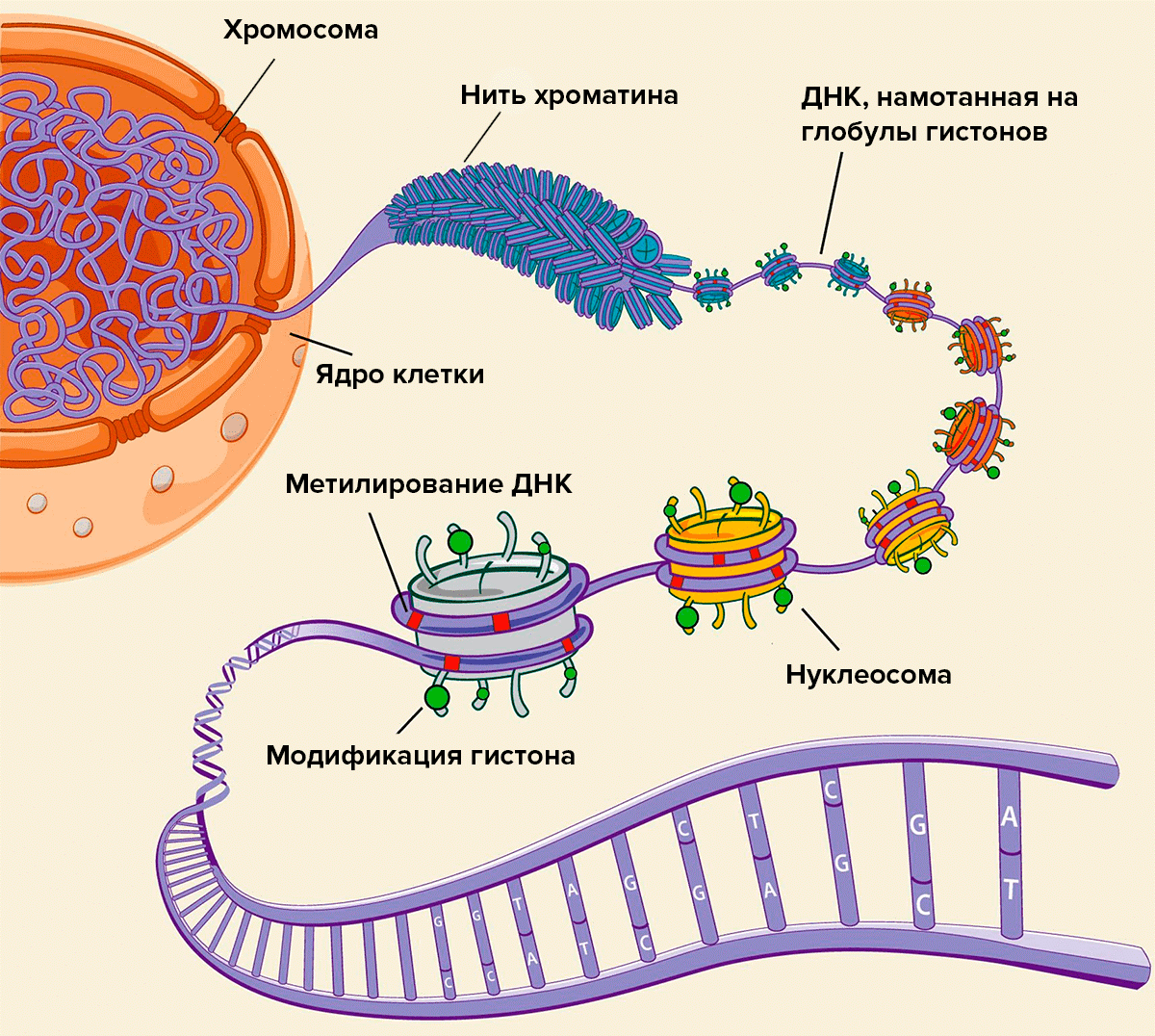
Рис. 1. На рисунке изображены уровни компактизации хроматина, состоящего из скрученных по спирали петель из ДНК.
По сути, эти генные вставки могут играть роль изначальных «настроечных» реперных «колок» (аналоги ключей настроек-натяжителей струн в гитарах). Разноимённые заряды стягивают, а одноимённые заряды разрыхляют, расталкивают нити ДНК, а значит степень упакованности, плотности, свёрнутости хроматина. Там, где одноимённых зарядов мало, а плотность упаковки усилена (аналог слабой натяжки струны – где как результат «струна не звучит»), там гены блокированы. Кроме того, на свойства мантии влияют множество внешних факторов, которые определяют статус эпигенома и судьбу клетки…
Концепция, что эпигеном и хроматин — это БИОКРИСТАЛЛ, а точнее устойчивый суперкристалл, то есть самоорганизующийся и самовосстанавливающийся, который после всех внешних воздействий и раскручивания и удвоения способен самостоятельно собраться, скрутиться в изначальное (автохтонное) состояние. Во время деления клетки происходит деспирализация (раскручивание) хроматина. Но при этом полного одномоментного раскручивания по всей длине не происходит, так как при частичном раскручивании (локальном) на этом же месте ДНК разъединяется на две стороны и обе из них тут же достраиваются в две новые хромосомы и тут же восстанавливают петлевую структуру тела хроматина.
Как происходит считывание информации. Этим занимаются ферменты – молекулярные машины, в первую очередь РНК-полимераза. Этот белковый комплекс раскручивает двойную спираль ДНК, словно молния на куртке, и считывает последовательность нуклеотидов, создавая на их основе молекулы РНК. Но «молния» работает только в том месте где нужно снять копию. Как она узнаёт это место? Очевидно, подсказку этому дают комплекс генов, принадлежащий данному уровню стволовости. Здесь гены деметилированы и активны.
Напомню, что особенностью суперкристаллов является то, что они обладают способностью после энтропийных факторов вернуть в исходное состояние все его ячейки, также и в хроматине - все его спирали. Нечто похожее восстановление (сжатие) происходит и у резинового жгута после его растяжения. Хроматину для этих свойств нужно поддерживать свои изначальные спирали аналогично суперпозициям в ячейках кристаллической решетки. Очевидно есть факторы, когда происходит отдаление, нарушение резонанса этих суперпозиций, при этом витки садятся не на свою исходную позицию. Это в свою очередь является фактором того, что ДНК хроматина удаляется от оптимального позиционирования своих программ. «Намотки», петли скручиваются не в такт, хаотически. Это может ослабить транскрипцию ряда программ или сделать их недоступными.
При этом процесс реализации невидимости программ при сенесценте не стохастичен, а именно заранее предопределён, коммитирован (на эпигеномно-прогениторном* уровне), что является особенностью для клеток животного типа. Причём сенесцентными клетки не рождаются, а становятся таковыми. Для реализации чего нужно сочетание нескольких факторов. Нужно запустить прогениторы на сенесцент. В этом случае происходит коммитация на уровне эпигенома, за счёт механизма «колок*» (действует типа ключа-натяжителя для струн на гитаре) или «реперных-“пробок”» - ограничителей на хроматине, которые и предопределяют в будущем его активность-пассивность на определённых участках.
Эпигенетическая регуляция – «редакторы» генетического текста. Эпигенетические механизмы выступают в роли редакторов и цензоров. Через эпигеном определяется какие гены будут «прочитаны», а какие останутся «закрытой книгой». Именно для этого работает метилирование ДНК и модификация гистонов – они маркируют различные участки генома, делая их доступными или недоступными для чтения. Наш код, таким образом, больше похож не на программный код, а на цензурированный текст, где большая часть информации скрыта под черными прямоугольниками редактуры. А разные типы клеток отличаются друг от друга именно тем, какие фрагменты текста остались для них доступными. Эпигенетические механизмы определяют, какие часть генетического кода «увидит» клетка. Условия для прочтения информации РНК-полимеразой создают не просто её возможности, а весь комплекс мантии и Биокристалла. Скопированная цепочка нуклеотидов сама по себе ничего не значит; значимость её смысла, назначения определяет Биокристалл. Он существует в четырёхмерном пространстве-времени. Он «знает» потребности организма и ткани. Он сенсор и редактор.
Но из этих позиций следует, что Биокристалл определяет! и редактирует текст, это означает, по сути, что он проявляет элементы сознания. Как он это делает? Внутри этого комплекса структуры геном-эпигеном ответа на этот вопрос не найдёшь. Очевидно, в его эту функцию входят ответы, которые находятся за их пределами. Выставляют метки метилирования многочисленные механизмы биокристалла, которые находятся не только внутри клетки, но и за её пределами: таковыми, очевидно, также являются механизмы метавоспаления* и прововоспаления.
Но в таком понимании это означает, что геном – это библиотека, которую считывают другие структуры. В тоже время есть мнение, что эпигеном – это операционная система, а ДНК – это аппаратное обеспечение, носитель информации, но сама по себе не выполняет действий. Эпигеном – это якобы набор инструкций. Но операционная система одновременно означает, что это набор программ, которые управляют всеми процессами в компьютерах. Но каждая программа – это всегда носитель информации, код, который управляет другими. Но в эпигеноме нет носителей программ. Очевидно эти программы записаны в геноме (тогда он тоже является операционной системой). Следовательно, программы записаны в геноме, а эпигеном только заводит тот или иной комплекс генов. Эпигеном не может быть носителем программ. Но из этого выходит, что программа – это не просто предписания для считывания, например, одного белка-фермента, а целая последовательность комплекс программ, которые осуществляются, считываются одновременно. Из этого можно предполагать, что некоторые программы генома могут сами себя индуцировать без воздействия со стороны эпигенома. Тогда становится понятным механизм метавоспалений, которые заводятся кейлонно, без эпигенетических влияний.
Итак, эпигеном не может быть опереционной системой, он лишь выполняет функции пускателя…
Редактирование метилированием скрывает текст. Считалось, что действие мета- и провоспаления направлены во вне, на становление фенотипов и модификатов. Но оказывается оно же направлено и во внутрь – на эпигеном, меняя его качественно, рисуя «рисунки» над геномом, а точнее над его активными или имеющими потенциал (скрытые) на активность точками (гены стволовости). Причем зоны над генами высшей степени стволовости в наибольшей степени метилированы и закрыты.
Но сознанием биокристалл не может обладать, он просто имеет предписания, коммитацию. Это аналогично тому как по внутренним правилам, предписаниям идут процессы морфогенеза, которые осуществляют трансформинг. Очевидно биокристалл тоже подчиняется неким предписаниям и проявляет свои особенности именно через обратную связь с геномом. Они относительно друг друга саморегулируются, самоопределяются. Некие состояния Биокристалла сопряжены с генами и в определённых состояниях кристалла эти гены подключаются. Причём, эти гены не просто подключаются, а обозначают где поставить «метки» эпигеному. Сам эпигеном это сделать не может. Так идёт эпигенетическая регулировка-редактирование, которая в итоге осуществляется через механизмы меток в виде метилирования. Таким образом, осуществляется взаимодействие между внешним и внутренним, между эпигеномом и геномом (геном предписывает где должны быть выставлены метки эпигенома), а эпигеном в свою очередь находится под контролем Биокристалла, а сам Биокристалл регулируется общим статусом гомеостазов, а последние определяют гормезис, то есть осуществляется сквозное саморегулирование. То есть, эпигеном и мантия тоже не самостоятельны, а строго сопряжены с геномом: одно читает другое и наоборот: они друг у друга под контролем. То есть «сознание» эпигенома записано на геноме. Таким образом, корректировка-редактирование и фиксирование (внешне это закреплённые фенотипы, модификаты) статуса каждой из сторон этой единой системы происходит относительно друг друга.
Таким образом, эти взаимоотношения эпигенома и генома являются «лазейкой», проходом для корректировки статуса клеток: ювентальные – сенесцентные.
Кстати, у растений тоже имеется мантия и биокристалл, но нет сенесцента и онкологии. Вся разница в том, что в растительных клетках регулировка их осуществляется только с одного направления – системного, то есть верхнего, через механизмы онтофизиса, поэтому даже если клетки и стареют, то это всего лишь ложное, навязанное старение. В свою очередь у клеток животного типа есть механизм коммитации сенесцента, то есть определяется автономно непосредственно в самих клетках, что является по сути истинным старением клеток.
Блокирование стволовых клеток высших потентностей. Особенность клеток животного типа:
- имеют особую глубокую коммитацию, реализуемую через механизмы провоспалений,
- обладают более высокой автономией.
Всё это обуславливает появление у них механизма сенесцента.
Это осуществляется у них через уровень метилирования, компактизацию-упаковку хроматина, но главное у них появляются провоспалительные механизмы регулировок, которые осуществляются на клеточном уровне, но которых нет у растений. Причём механизмы блокирования стволовости здесь иные и блокировка происходит изначально на этапе эмбриогенеза.
Причём колок-натяжителей (аналог ключей настройщиков звучания для каждой струны) существует столько сколько в эпигенезе отключается, глушится степеней потентности стволовых клеток. Первую очередь глушатся: тотипотентные, плюрипотентные, мультипотентные, унипотентные… Очевидно, это те, которые связаны с морфогенезом. Подавление ювентальности и переход на сенесцент очевидно в большей степени зависит от прогениторного типа (концевых по степени потентности) стволовых клеток. С прогениторами связан эпигенез и сенесцент.
Итак, очередность «глушения» стволовости у различной степени потентности стволовых клеток различная:
- наиболее высокой степени потентности клетки глушатся на много раньше, а тотипотентные – на стадии эмбриогенеза,
- последним глушение происходит в системе «эпигенома ↔ прогениторов».
Как происходит это разновременная блокировка? Очевидно на это сказываются эдафотопы и коммитации.
Итак, некоторые программы, отвечающие за уровень потентности стволовых клеток, «запечатываются» на ранних этапах Онтодианной Линии Жизни, допуск к ним закрыт, они становятся «невидимыми». Это ограничивает-исключает различные уровни глубокой регенерации, ослабляет потенциал ювентализации в пользу сенесцента, а также отключает репарации теломер. «Пробки»-«колки» (работают подобно вентилю: закрыто-открыто) на эпигеноме, по сути играют роль первичной «надпрограммы», а точнее указателя для прогениторов. Работа прогениторов в первую очередь настроена на реагирование, подстраивание на внешние сигналы. Так что степень доминантности сенесцент/ювентальность реализуется на уровне прогениторов.
Действуют эти колки-реперы подобно теломеразе, которая отвечает за восстановление теломер. От состояния натяжимости ДНК на её концах зависит баланс натяжённости и вдоль других участков тела хромосомы. Натяжимость, очевидно, также сказывается на степени поляризации, величине электростатических зарядов на ДНК. Но у «колок» есть аналоги «теломеразы», которые регулируют величину-длину нуклеотидных вставок (удлиняя или укорачивая эти вставки), тем самым снимая излишнюю натяжку или наоборот её увеличивая в данном месте, аналогичных как у теломер. Эти вставки могут регулировать степень натяжимости хромосомы в данном месте. Длина этих вставок может, также, как и теломеры, глушить работу ряда генов. Но если теломеры находятся на концах хромосом, то вставки (инсертионемы) находятся возле комплекса генов, отвечающих за активность программ, обеспечивающих различную степень потентности стволовых клеток.
Клетки становятся с усечёнными возможностями, отсутствует полный цикл репараций. Это в свою очередь открывает возможность для появления в последующем эпигенотипов и делает возможным сенесцент клеток на определённых этапах развития, причём ускоренно в определённых типах ткани. Автор выдвинул концепт, что клетки целенаправленно используют этот механизм «глушения» для запуска сенесцента и используют его в целях эпигенеза и смены фенотипов. Возможно с помощью сенесцента они останавливают непрерывный поток роста клеток, там, где этого делать нельзя, в частности в высокодифференцированных клетках? Это уже путь фенотипизации (в отличие от морфотипов) и обеспечивает становление этапов и продвижение по онтодиане*. Таким образом, системы клеток и тканей активно применяют регулировки сенесцентом, управляют им, наводят его локально через макрофаги в нужных системах. Если это так, то можно утверждать, что нет специфического гена (-ов) вызывающего тотальный сенесцент во всех клетках? Клетки животных изначально предрасположены к управляемому сенесценту. Кроме естественного всеобщего сенесцента, предписанного лимитом Хейфлика, существуют и механизмы запуска ускоренного и направленного на местах сенесцента. Возможно тотальный лимит Хейфлика реализуется на уровне теломер, а локальный сенесцент на уровне инсертионем*. Этому пути «глушения» могут способствовать банальные механизмы клеточного неспецифического стресса УКАС, ОАС, SASP, адальтус, метилом, сенилит и др. Общее у всех у них, то что они создают новые «эдафотопы» (среду, степень метилирования или…), которые влияют на качество хроматина.
В стрессовых условиях, когда клетки подвергаются воздействию различных факторов и мутагенов, анионная форма нуклеотидов увеличивается. Анионная форма нуклеотидов – это состояние, в котором азотистые основания (аденин, гуанин, цитозин, тимин) периодически меняют форму, переходя в таутомерные и анионные состояния. Это может происходить на тысячные доли секунды. Это повышает риск не только ошибок, но и степень метилирования и электростатику… Ковалентный остов нуклеотидов состоит из монотонно чередующихся фосфатных и пентозных групп, которые несут отрицательный заряд.
Эдафотопы (разные среды вокруг эпигенома) обуславливают появление новых эпигенотипов. Блок «эдафотоп-эпигенотип» срабатывает возле определённых стволовых клеток, пробуждая или тормозя их активность, то есть действует локально, где и образуются метки. В этом случае эпигенез является как продолжением, так и следствием «глушения» хроматина и активности паттернов над стволовыми клетками. Отсутствие поддержки со стороны стволовых клеток является фактором для ослабления-нарушения соподчинения ядра и митохондрий в рядовых клетках-сателитах, всё заканчивается митохондриальной недостаточностью. При этом хроматин особенно чувствителен в реперных точках-«пробках», а также это может ускорить недокручивание (ослаблена степень натяжимости ДНК) в местах окончания хромосом, которые завершаются теломерами. Здесь же на скрутках находятся и гены теломеразы (восстанавливает теломеры – защитные колпачки) и реализуется эпигенез? Тогда механизмы эпигенеза должны совпадать с механизмами укорочения теломер. Это означает что эпигенез первичен, а процессы митохондриальной недостаточности и сенесцента вторичны.
Удивительно, но феномена теломеразной недостаточности на хроматине или лимита Хейфлика нет у растений. Они обладают идеальным самовосстанавливающимся биокристаллом хроматина. Почему? Степеней свободы к автохтонности хроматина у них намного больше? В тоже время у растений тоже есть эпигеномные аберрации и фенотипы…
Итак, можно предполагать, что удаление от оптимального позиционирования своих программ ДНК у хроматина происходит именно на концах скруток (спиралей) и инсертионемах, где и происходит «глушение» программ.
3D карты хроматина в качестве подтверждения значимости скруток ДНК для нормальной или ненормальной её активности [1]. Удалось создать наиболее детальные и всесторонние карты того, как ДНК сворачивается, образует петли, домены и меняет своё положение внутри живых клеток в трёх измерениях и во времени. Эти карты раскрывают ранее скрытый слой генетического контроля: не только сама последовательность нуклеотидов, но и пространственное расположение генов и регуляторных элементов определяет, какие гены будут активны, а какие – молчать.
Оказалось, что гены и регуляторные участки, которые находятся далеко друг от друга в линейной последовательности, могут оказаться в непосредственной близости в пространстве благодаря петлям (виткам) и складкам. Именно эти контакты позволяют «дальним» энхансерам* (усилители) активировать нужные гены или, наоборот, изолировать их от влияния посторонних регуляторов. Выяснилось, что без 3D взаимодействий ДНК не сможет нормально функционировать.
Более 140000 хроматиновых петель (витков) идентифицировано в каждом типе клеток (эмбриональные, стволовые и фибробласты). Выявлены якорные элементы, которые удерживают эти петли. Описаны хромосомные домены и их положение внутри ядра. Показаны различия в организации генома между отдельными клетками одного типа. Продемонстрировано, как структура меняется в процессе роста, деления и функционирования клетки.
Сенесцент это проявление свойств Биокристалла – такой вывод можно сделать на основании выше перечисленных утверждений.
Также хроматин подобен пьезокристаллу, когда любое воздействие на него формирует электрический заряд или под действием электрического напряжения кристалл меняет форму, например, скручивается. Пьезоэлектрический эффект – результат поляризации диэлектрика под действием различных причин. При этом происходит смещение связанных зарядов или поворот диэлектрических диполей. Очевидно роль таких диполей играют гистоны в гистоновой рубашке. Гистоны – ядерные белки, которые входят в состав хроматина и содержат положительно заряженные аминокислоты – лизин и аргинин, что позволяет им взаимодействовать с отрицательно заряженными фосфатными группами ДНК и нейтрализовать их заряд. Гистоны обеспечивают компактную упаковку ДНК. Гистоны действуют как катушки (нуклеосомы), вокруг которых наматывается ДНК (см. рис. 1). Гистоны участвуют в эпигенетической регуляции транскрипции, репликации и репарации. Они же участники и модификаций. Это же может происходить и под действием различных магнитных полей и др. сил. Важной особенностью такого заряженного хроматина является автоподдержание баланса в электростатической намагниченности в каждом из звеньев, образующихся при спиральном скручивании двухцепочной нити ДНК. При этом любое локальное изменение влияет на общее механическое напряжение во всей цепочке и нарушает транскрипцию. Каждая функциональная скрутка (звено) должна лечь в своё ложе (гнездо) и образовать свой функциональный паттерн (гистоновый код).
Эпигенез сенесцента и онкологии ломает механизм нормального скручивания и транскрипции. При этом паттерны ложатся не в своё ложе (гнездо) и начинается путаница в скрутках, образование узлов в хроматине с потерей зарядов на некоторых паттернах, что делает их невидимыми (сайленсинг*) или наоборот. Очевидно каждая скрутка обладает зарядом как на обкладках конденсатра, что поддерживает хромосому как в подвешенном состоянии (невесомости) на плаву в электромагнитном поле. При ослаблении зарядов хромосома сдвигается от центра ближе к оболочке ядра («выпадает в осадок»). Но в любом случае (сенесцент или онкология) нарушена энергетика и разность потенциалов, причём в разной степени. Сенесцент – следствие гистоновых модификаций. Онкология – тоже является проблемой гистоновых модификаций, а не мутаций.
Автор предлагает концепцию, что ювентальные или сенесцентные клетки имеют каждая свой эпигенетический статус (эпигенотип) и свою эпигенетическую среду – эдафотоп. Каждый эдафотоп - это, по сути, ландшафт-ниша, которую определяют, как свои внутриклеточные SASP или UASP, так и выделения на уровне всего организма, включая продукты ОАС, общий метаболом, гормональный фон и др… В совокупности они могут обусловливать определённый статус преобладания процессов метилирования или ацетилирования на гистоновой рубашке и в определённой системе её паттернов (эпигенотип). Каждый эпигенотип может существовать в условиях своего эдафотопа или другими словами каждый эдафотоп имеет свой эпигенотип. Каждый эпигенотип имеет свой фенотип (модификат).
Почему у МF2 могут быть такие разные фенотипы клеток? В обоих случаях отмечается митохондриальная дисфункция, при которой происходит падение электропотенциалов на мембранах клетки, включая митохондрии, ЭндоПлазматическийРетикулум, ядра. Очевидно различия связаны с разной величиной падения этих потенциалов: у сенесцентных клеток он снижается с -70 мВ до -50 мВ, а у онкологических – до -40 мВ. Этого достаточно чтобы изменить состояние эпигенома. Различная степень дисфункции!? Различный запуск опосредованных эпигеномно экспрессий, например, в ядре клетки ген р53? Тогда следует говорить о дисфункциональных подтипах среди типов MitFen2, то есть о MitFen2/Dis1 (сенесцентный) и MitFen2/Dis2 (раковый). В обоих случаях и раковые, и сенесцентные клетки связаны с недовыработкой потенциалов, но параметры недовыработки существенно различаются, что и запускает разные эпигеномные паттерны и сдвиги сказываются на работе разных генов? Сенесцент и онкология – два противоположных крыла одной системы репликаций. Клетка как бы упускает из рук вожжи самоуправления и согласовывания их с потребностями целого. Это означает выход за пределы гомеостазов, где невозможно ауторегулировка клетки. Вместо автозапуска механизма возврата к гомеостазу (центропии*) запускаются механизмы энтропии, усугубления проблемы, то есть в противоположном направлении и раскачивания клетки в неуправляемые процессы.
Термины
— Белки теплового шока — БТШ – биологические маркеры клеточного стресса, экспрессия которых обычно усиливается при повышении температуры и др. факоторах.
— Геронтос — общее старение регулируется одновременно двумя рычагами на двух этажах: на нижнем уровне фенотипа клеток (сенесцент) и на верхнем уровне всего организма в виде онтофизисных фенотипов (сенилит). — Термин предложил Г. А. Гарбузов.
— Инсертионемы (-а) — дословно означает вставка и подразумевает вставки в хромосомах, которые регулируют её натяжку и тем самым обеспечивают видимость или невидимость стволовых клеток. Действуют и регулируются подобно теломерам, которые регулируются активностью фермента теломераза. От степени метилирования и натяжки хромосом теломераза может быть заблокирована. У инсертионем имеются свои «теломеразы». – Термин предложил Гарбузов Г. А.
— Конформационные белки — меняющие свою форму в пространстве и регулируются различными факторами.
— Метавоспаление — воспаление происходящее изначально в некоторых типах иммунных клеток (кейлонные механизмы) и которое затем передаётся другим клеткам в виде провоспалений. – Термин предложил Гарбузов Г. А.
— Прогениторные клетки — стволовые клетки самого последнего крайнего типа стволовости. Предполагается, что они больше всего взаимодействуют с эпигеномом. Подавление ювентальности и переход на сенесцент очевидно в большей степени зависит от прогениторного типа (концевых по степени потентности) стволовых клеток. С прогениторами связан эпигенез и сенесцент.
— Рендерировать, рендеринг — дословно «визуализация», процесс преобразования трехмерной модели в двухмерное изображение.
— Реювентализация — омоложение клеток, снятия с них фенотипа сенесцента с помощью механизмов ювентального реверса. – Термин предложил Г.А. Гарбузов.
— Сайленсинг — подавление экспрессии генов. Это общий термин, описывающий эпигенетический процесс регуляции генов. При этом последовательность нуклеотидов не изменяется, а лишь прекращается экспрессия соответствующего гена.
— Сенилит — старение на общеорганизменном уровне.
— Центропия — процессы-механизмы противостоящие энтропии, то есть обратное действие - Термин предложил Г.А. Гарбузов.
— Энхансер — дословно «усилитель» - участок ДНК, который после связывания с ним факторов транскрипции стимулирует транскрипцию основных промоторов гена (-ов).
— Ювентос — состояние организма, когда в нём сочетаются ювентальность (клеточный уровень) и ювенильность (общеорганизменный уровень). – Термин предложил Г.А. Гарбузов. Противоположное состояние Геронтосу = Сенесцент + Сенилит.
Литература
Книги автора
 ЭПИФИЗ — ВЕРХОВНЫЙ ДРАЙВЕР ОНТОГЕНЕЗА. КЛЮЧ К МОЛОДОСТИ И СТАРЕНИЮ
ЭПИФИЗ — ВЕРХОВНЫЙ ДРАЙВЕР ОНТОГЕНЕЗА. КЛЮЧ К МОЛОДОСТИ И СТАРЕНИЮ
 ВЗЛОМ ИММУННЫХ МЕХАНИЗМОВ СТАРЕНИЯ — ПУТЬ К ЮНОСТИ И ЗДОРОВЬЮ
ВЗЛОМ ИММУННЫХ МЕХАНИЗМОВ СТАРЕНИЯ — ПУТЬ К ЮНОСТИ И ЗДОРОВЬЮ
 ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ. КАК МАГНИЙ И ОКСИД АЗОТА ПРОТИВОСТОЯТ СТАРЕНИЮ
ЗДОРОВЬЕ, МОЛОДОСТЬ, ДОЛГОЛЕТИЕ. КАК МАГНИЙ И ОКСИД АЗОТА ПРОТИВОСТОЯТ СТАРЕНИЮ
Глубокий анализ существующих причин старения позволил автору создать свою единую теорию «биологии старения», которая строится на интеграции процессов сенесцента (на уровне клеток) и сенилита (на системном уровне). Показана решающая роль магния и оксида азота в регулировке на клеточном уровне сенесцента. Именно сенесцент (нарушение гомеостазов) определяет механизм сенилита (нарушение гормезиса и витаукта), а с ними и возрастзависимых «болезней», которые на самом деле не болезни, а естественные анизомалии, то есть механизмы реализации старения. Только правильное понимание сути природы старения позволит выйти на правильные пути преодоления возрастных болезней и продления жизни человека через механизмы неотении (продлённой молодости), чему посвящена следующая книга автора «Биология молодости».
 САМОУНИЧТОЖЕНИЕ РАКОВЫХ ОПУХОЛЕЙ. ФЕНОМЕН АПОПТОЗА
САМОУНИЧТОЖЕНИЕ РАКОВЫХ ОПУХОЛЕЙ. ФЕНОМЕН АПОПТОЗА
Онкология. Можно ли её вылечить безвредными способами? Автор впервые показывает, что можно и нужно! Для этого нужно запустить механизмы избирательного самоуничтожения раковых клеток в организме на основе клеточных программ апоптоза и аутофагии, которые нужно адресно открыть. Предложена Комплексная Программа из противоположно направленных методов, вычлененных в две фазы и направленных на расшатывание и перебалансировку основных жизнеобеспечивающих механизмов клеток, в т. ч. антиоксидантной « прооксидантной, провоспалительной « антивоспалительной и др. У онкоклеток коридор возможностей самозащиты, их гормезис намного уже, чем у нормальных клеток, у которых Жизненная Сила, Витаукт, намного выше. Такая разность возможностей позволяет создавать особые условия, когда становится возможным запуск механизмов самовыбраковки онкоклеток.
 БИОЛОГИЯ МОЛОДОСТИ – электронная рукопись 2026 г
БИОЛОГИЯ МОЛОДОСТИ – электронная рукопись 2026 г
Практически все авторы научно-популярных работ по анти-эйдж теме рассматривают всевозможные методы сдерживания старения и продления жизни, здоровья… Автор книги впервые показывает, что это не эффективно, так как не убирает суть проблемы и связывает эту тему через продление молодости, которая и есть фундамент для всего остального. Без продления молодости невозможно радикального решения всех остальных проблем. Автор подробно анализирует биологию молодости, объясняет почему она уходит. Важнейшими для запуска старения являются механизмы неспецифического иммунитета, а также старение клеток (сенесцент). Изучен весь мировой опыт и знания по продлению молодости на всех уровнях организма, что автоматически отодвинет все «болезни» старости - анизомалии.
Книгу или Программу-консультацию по заболеваниям Вы можете приобрести по нашему адресу.
Вы можете обратиться с вопросами или за консультацией к Гарбузову Геннадию Алексеевичу на сайте garbuzov.org, по адресу: vitauct@yandex.ru или по телефону 8 (928) 239-13-64.

Похожие статьи
Биолог, дипломированный фитотерапевт, нутрициолог, кандидат биологических наук
Стаж 40 лет
Подробнее обо мне